
Hippo ;����������
Hippo ;���ǹ�Ӭ�Ͳ��鶯���и߶ȱ��صļ����ź�;����ͨ�����ڹؼ��л�������������ѧ���̣�����ϸ����ֳ�����ֻ���ϸ�����˾��������ٴ�С����֯��̬���� Hippo ;�����ź��쳣Ҳ�漰���ֲ�����������֢�����ߵȡ�
YAP/TAZ �� Hippo ͨ·��������Ҫ��ת¼����������������ϸ���ʺ�ϸ����֮�䴩�� (Shuttle)��YAP/TAZ ������ Hippo ;������Ŧ�����εĶ����źŷ���ͨ��ֱ�ӻ��������� YAP/TAZ����Ҫ������ YAP/TAZ �Ķ�λ�������� YAP/TAZ ������ϸ���ʻ��ǽ��к˶�λ (��λ��ϸ���ˣ������Ӧ��ת¼��������ã��Ե��ذл���ı���)��
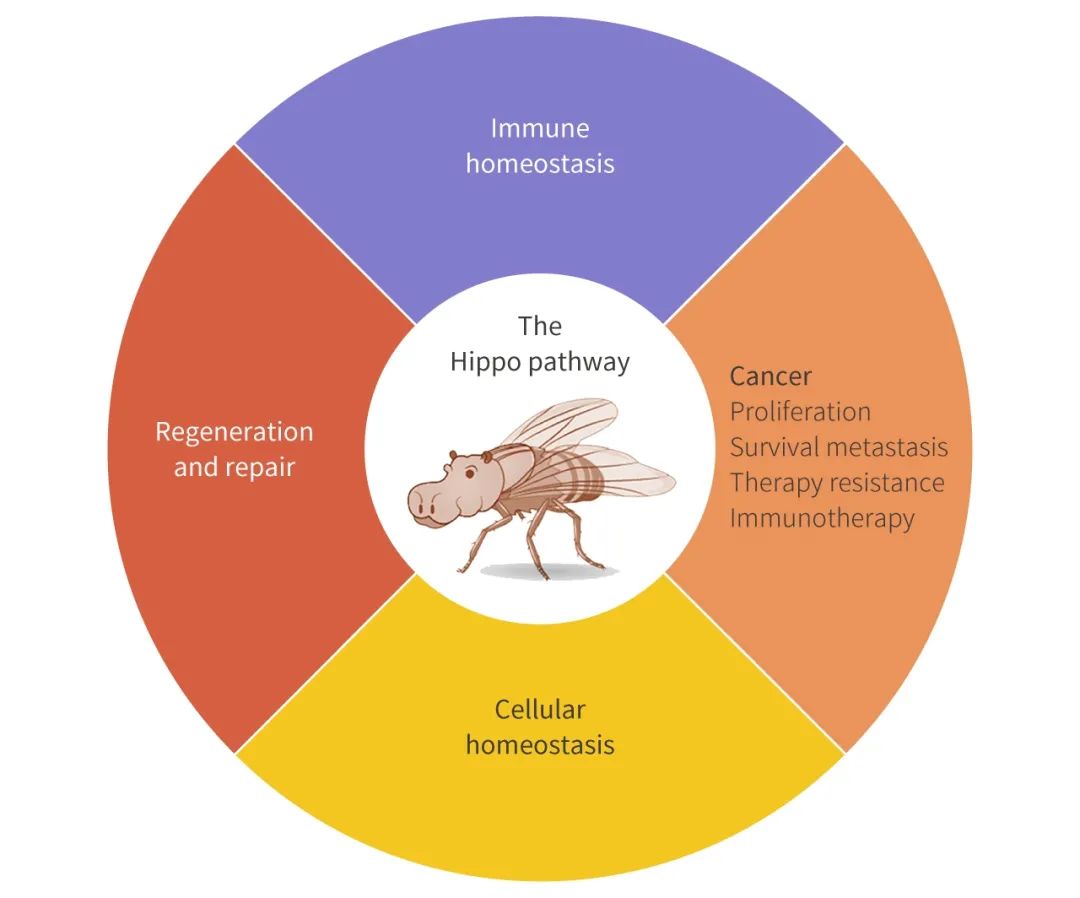
ͼ 1. Hippo ;�����������������[5]
Ŀǰ������ Hippo ;������־��� 30 ���֣������� Hippo ;������Ҫ��ֱ��Ͷ�Ӧ��ͨ·ͼ��
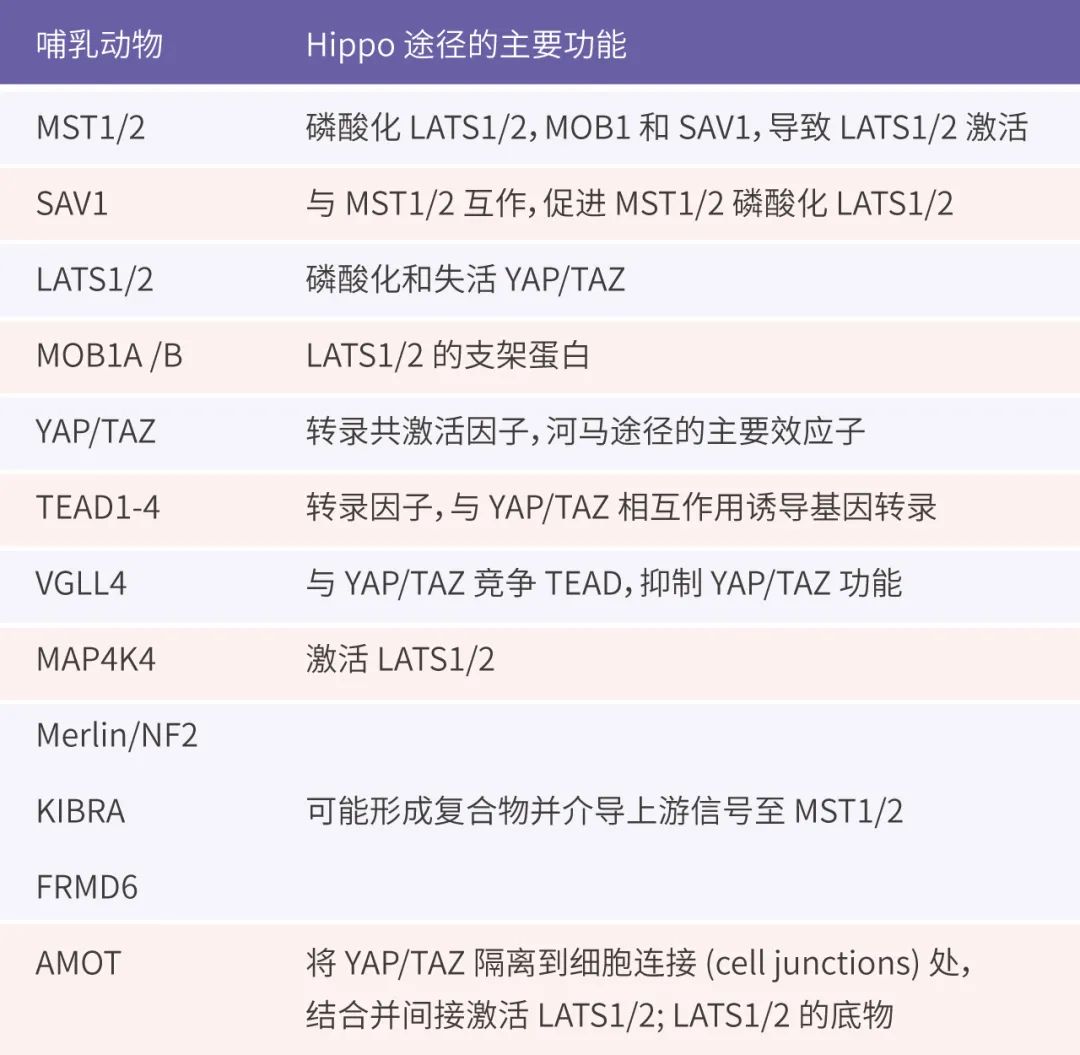
Hippo ;����Ҫ����빦��[6]
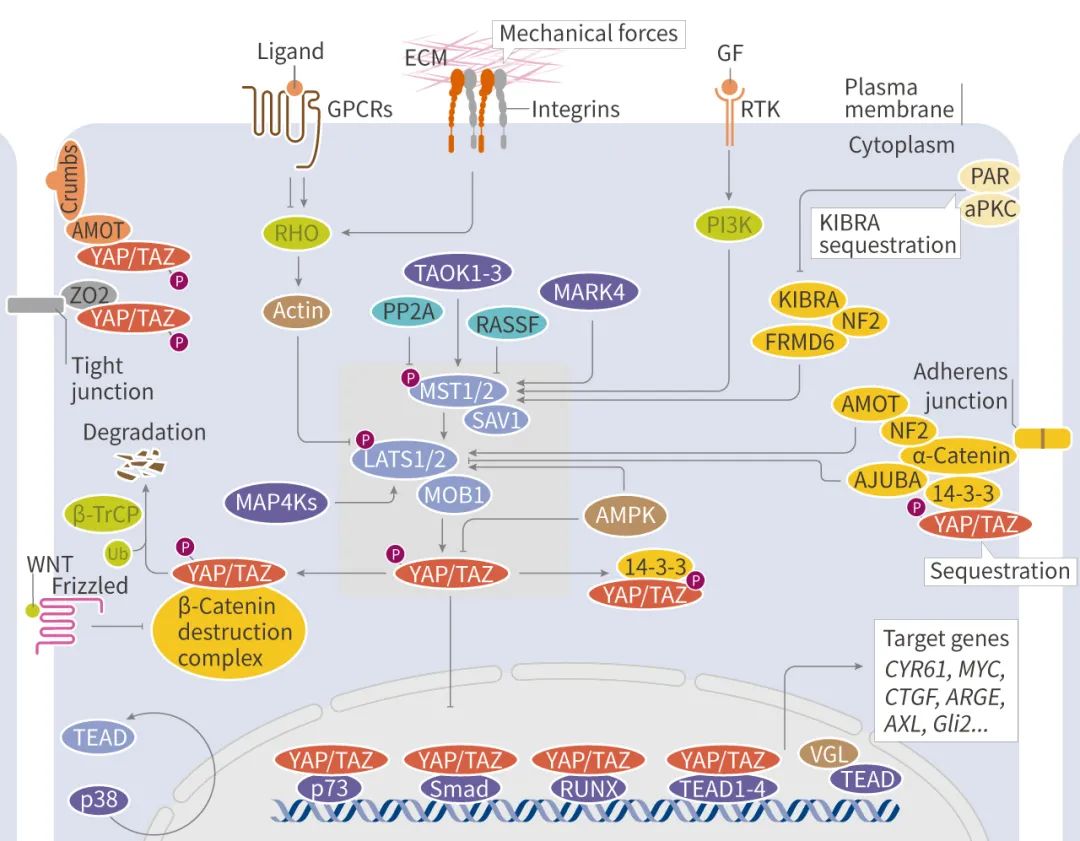
�˳��ؽ������㣬����ͼ���Dz���������ӵģ�

Ϊ�˸��õ��������ͨ·��С��������������������¿�ͼ��ϰ�ߣ��� Hippo ;���ĺ��ġ���MST-LATS ��ø������Ӧ�������������ź����������ת¼���ء�
�� ��ø������Ӧ
MST-LATS ��ø����;����һ�鼤ø (MST1/MST2, LATS1/LATS2) ���νӵ���SAV1��MOB1 ��ɡ����������ź�ͨ������ MST-LATS ��ø������Ӧ�����������°�ת¼���������� YAP �� TAZ �Ķ�λ������Ϊ���� Hippo ͨ·��ʱ���� SAV1 �γɸ������ MST1/2 ��ø���ữ������ LATS1/2-MOB1 ������ �� ��� LATS1/2-MOB1 Ȼ�����ữ YAP/TAZ �� ���ữ�� YAP/TAZ �� 14-3-3 ���ױ�����ϸ�����л�ͨ�� ��-Trcp E3 ����ø�鵼���⣬���� YAP/TAZ �ĺ����� �� �µ����ΰбꡣ�� Hippo ͨ·�ر�ʱ��YAP/TAZ δ�����ữ������λ��ϸ�����У��� DNA ���ת¼���ӽ�ϣ��ٽ��л���ı��
�� ���ζ����ź�����
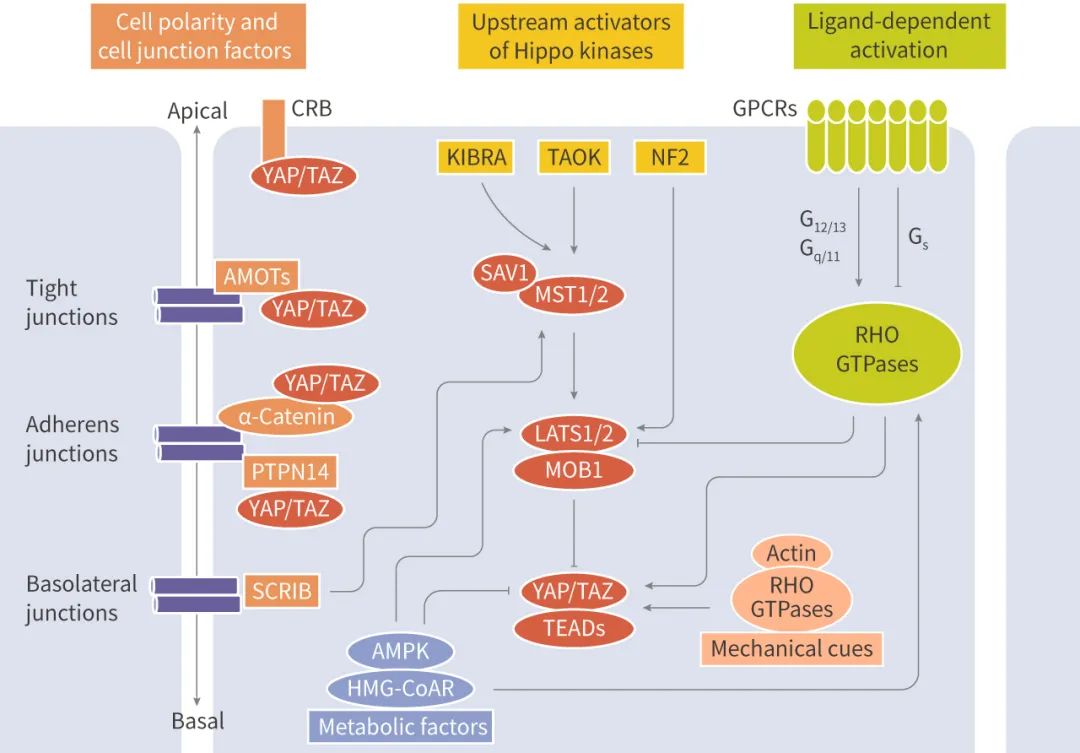
MST-LATS ��ø���������ữ�¼��ܵ����ζ����źŵ��ڣ����� ��
1) �鵼 Hippo ���ļ�ø������������纬 FERM �ṹ��� NF2 ���ס�KIBRA ���ס�TAO ��ø (TAOK);
2) ϸ�����Ը���������ӵ��������Ժ�ճ�����ף��� ��-catenin��E-cadherin��CRUMBS �� SCRIBBLE ������ɽ� YAP/TAZ ������ϸ������;
3) G ����ż������ (GPCR) �Ϳ��������������� GPCR ��������� LATS1/2 ȡ���� G�� �������ͣ��� Run Fan et al. ֤ʵ����ϸ�� EGFR ͨ������ PI3K �� PDK1 ���� Hippo ;�������� YAP �ļ���;
4) �� Rho �ͼ������ۺϵ��ڵĻ�еӦ���ı仯��Ҳ�ɵ��� YAP/TAZ �˶�λ;
5) ϸ��Ӧ���ź�������Ӧ����������Ӧ����ȱ���ȶ� YAP �� TAZ Ҳ�е��ڡ���ϸ������Ӧ���ܹ����� AMPK���Ӷ�ͨ��ֱ�����ữ���ȶ��������ӵ��� AMOTL1 ������ YAP��
����������Щ�źŴ������������������ֵ���;������ Hippo �ź�ת������ MAP4Ks �����ữ������ LATS1/LATS2��AMOT ���嵰��ֱ���� YAP1 �� TAZ ��ϣ��ڽ������Ӵ��������Dz��������ǵ�ת¼���ԣ�����ͨ�� LATS1/2 ;����ӵ��� YAP/TAZ��PP2A �� MST1/MST2 ��ø����ò�ʹ��ȥ���ữ������ YAP/TAZ ���
����;�����뾭�� Wnt ;�� (Wnt �ź�ͨ·�ܸ��ӣ�) �� PI3K ;���ȵ�ת¼���� ��-catenin ����ã��Ӷ����䱣����ϸ�����С�
�� ����ת¼����
YAP/TAZ ͨ�������� DNA ת¼�����������ʶ��һЩ˳ʽ����Ԫ�������ڰл������鵼�� Hippo ͨ·����Ҫ������غ�����ѧ���������У�TEAD1-4 �� YAP/TAZ ��Ҫ�����ת¼���ӣ�YAP/TAZ-TEAD �����������ϸ����ֳ�������ʹ��İл��������ӽ��, ���� CYR61��CTGF��AREG��MYC��Gli2��AXL��Bcl2 �ȡ��� VGLL ���嵰�� VGLL4 ���� YAP �����Խ�� TEADs��Ӱ�� TEAD �л���ı��
���⣬YAP/TAZ �������� DNA ���ת¼��������ã��� p73��ERBB4��EGR-1��RUNXs��SMADs �ȡ�
�� ���� Hippo ;������
�����о����� Hippo ͨ·�ļ����������ּ����йأ��� MST1/2 ������Ӧ���йأ�LATS1/2 �방֢�������йأ��� YAP/TAZ ����������������ձ鼤��ĸ߶���ص�ת¼�����������ڶ��ְ�֢�о���������ٽ������ķ�����������ת�ƺ���ҩ�ԡ���ˣ����� Hippo ;��������������
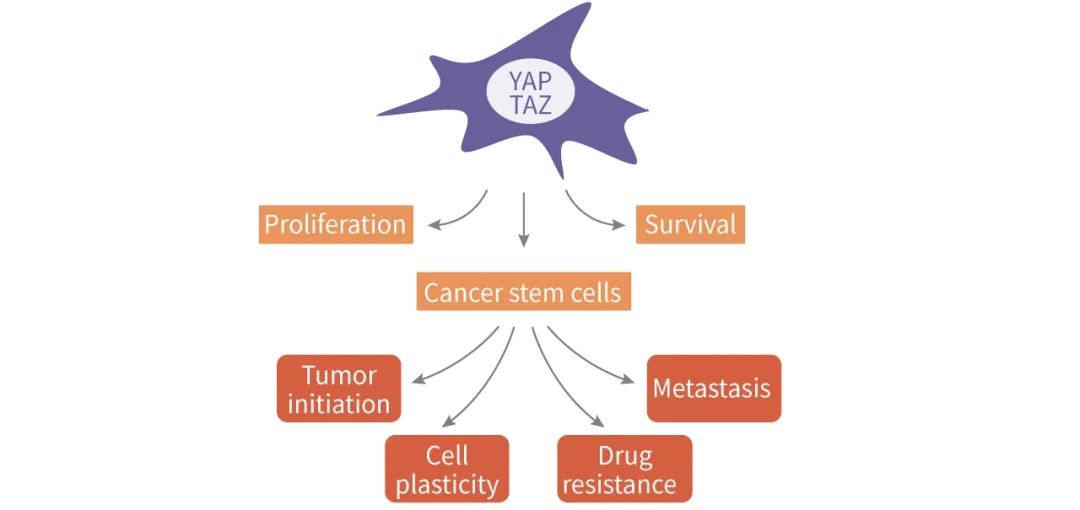
ͼ 4. YAP/TAZ �ڰ�ϸ���е�����[17]
1������ YAP/TAZ-TEAD ������
���� YAP �� TAZ ����Ľṹ���ʣ�ֱ�Ӱ��� YAP/TAZ ���ײ�����������ˣ����� YAP/TAZ-TEAD ���������һ�����õIJ��ԡ����� TAZ/YAP-TEAD �����Ƽ��� Verteporfin��Flufenamic acid��TED-347 �� VGLL4 ģ���� (VGLL4-mimicking peptide)��
2��������ļ�ø
���� MST1 �� MST2 �����Ƽ��� XMU-MP-1���Լ���ֹ MST/LATS ���ữ�����Ƽ� C19 (EMT inhibitor-1) �ȡ�
3�����������ź�
�����г��� Hippo ;���ܵ����������źŵļ�����Ҳ���������η��ӡ������ϸ���������� (�� GPCRs��EGFR)������ϸ���ڼ�ø (�� PI3K��MEK)���������� (Actin) �����Լ�����Ӧ�����ڡ�
4�������� YAP/TAZ ���ص��°�����
��ͨ����������ת¼�б��� BCL-xL��FOXM1��TG2 �� COX-2���Կ� YAP/TAZ �鵼���°��ԡ�
|
���� YAP/TAZ-TEAD ������ |
| Verteporfin YAP ���Ƽ������ƻ� YAP-TEAD ����ã��յ�ϸ������ (apoptosis)�� |
| Flufenamic acid �� TEAD2 YBD ������ڴ���ϣ����� TEAD ���ܺ� TEAD-YAP �����Ĺ��̣���ϸ��Ǩ�ƺ���ֳ�� |
|
Dihydrexidine |
| ���������ź� |
| Wortmannin
������ģ�ѡ���� PI3K ���Ƽ���������ֹ YAP ����ϸ������ |
| LY294002
���� PI3K ���Ƽ���������ֹ YAP ����ϸ���ˡ� |
| Cytochalasin D
��Ч�ļ������ۺ����Ƽ����������� YAP/TAZ �ĺ���λ�� |
|
ѡ���ԵķǼ��⼡�� II (NMII) ���Ƽ����������� YAP/YAZ ����λ�� |
| ML-7
���ǰ�����������Ƽ���������ø���������� YAP/TAZ �ĺ���λ�� |
|
���������Ҳ�� AMPK ����������յ� YAP/YAZ ���ữ�� |
| ������ YAP/TAZ ���ص��°����� |
| Celecoxib
ѡ���Ե� COX-2 ���Ƽ�����ͨ������ COX-2���Ӷ��Կ� YAP/TAZ �鵼���°��ԡ� |
| Thiostrepton
��Ȼ�Ļ�״���Ŀ����أ������� FOXM1 ��ת¼���ԡ�FOXM1 �� YAP/TEAD �������ϡ�YAP/TEAD/FOXM1 �������ڿ���ϸ�����ڵĻ�����������Ͽ��ܻ�Ӱ��ϸ����ֳ�� |
�ڹ�ȥ��ʮ���У����ǴӶ����˵� Hippo ;�����о���֤ʵ�� Hippo �ź��ڻ����е�������ã���ʧ���������֢�йء���ʵ���ϣ����Ƕ� Hippo ;������ʶ��Ȼ����ȫ�档С��������Ҳֻ�ǹ��п��������½����� Hippo ;�������ε���Ҫ��֣������� Hippo ;������������;�� (�� Wnt��TGF-�� ��) �����ϵ������ƪ��ԭ��û��չ�������������ʣ�Hippo ;�������ɼ�������ɡ���һ�л���Ҫ���Ǹ�������о� Hippo ;����Ϊ���������ṩ����Ч�IJ��ԡ�
���
YAP/TAZ��Yes-associated protein/Trascriptional coactivator with PDZ-binding motif
�����
�� �»��鿴��������
1. Lu Y, et al. Phase separation of TAZ compartmentalizes the transcription machinery to promote gene expression. Nat Cell Biol. 2020; 22(4): 453-464.2. Cai D, et al. Phase separation of YAP reorganizes genome topology for long-term YAP target gene expression. Nat Cell Biol. 2019; 21(12): 1578-1589.
3. Song K, et al. Yes-Associated Protein in Kupffer Cells Enhances the Production of Proinflammatory Cytokines and Promotes the Development of Nonalcoholic Steatohepatitis. Hepatology. 2020; 72(1): 72-87.
4. Xu L, et al. SHANK2 is a frequently amplified oncogene with evolutionarily conserved roles in regulating Hippo signaling [published online ahead of print.Protein Cell. 2020 Jul 13.
5. Calses PC, et al. Hippo Pathway in Cancer: Aberrant Regulation and Therapeutic Opportunities. Trends Cancer. 2019; 5(5): 297-307.
6. Yu FX, et al. Hippo Pathway in Organ Size Control, Tissue Homeostasis, and Cancer. Cell. 2015; 163(4): 811-828.
7. Moya IM, et al. Hippo-YAP/TAZ signalling in organ regeneration and regenerative medicine. Nat Rev Mol Cell Biol. 2019; 20(4): 211-226.
8. Bae SJ, et al. Activation mechanisms of the Hippo kinase signaling cascade. Biosci Rep. 2018; 38(4): BSR20171469. Published 2018 Aug 29.
9. Del Re DP. The hippo signaling pathway: implications for heart regeneration and disease. Clin Transl Med. 2014 Dec;3(1):27.
10. Sebio A, et al. Molecular Pathways: Hippo Signaling, a Critical Tumor Suppressor. Clin Cancer Res. 2015; 21(22): 5002-5007.
11. Run Fan, et al. Regulation of Hippo pathway by mitogenic growthfactors via phosphoinositide 3-kinase and phosphoinositide-dependent kinase-1. Proc Natl Acad Sci U S A. 2013 Feb 12;110(7): 2569�C2574.
12. Ma S, et al. The Hippo Pathway: Biology and Pathophysiology. Annu Rev Biochem. 2019; 88: 577-604.
13. Marcello Maugeri-Sacc��, et al. The Hippo pathway in normal development and cancer. Pharmacol Ther. 2018Jun;186:60-72.
14. Boopathy GTK, et al. Role of Hippo Pathway-YAP/TAZ Signaling in Angiogenesis. Front Cell Dev Biol. 2019; 7: 49. Published 2019 Apr 10.
15. Yu FX, et al. The Hippo pathway: regulators and regulations. Genes Dev. 2013; 27(4): 355-371.
16. Kim MK, et al. DNA binding partners of YAP/TAZ. BMB Rep. 2018; 51(3): 126-133.
17. Zanconato F, et al. YAP/TAZ at the Roots of Cancer. Cancer Cell. 2016; 29(6): 783-803.
18. Dey A, et al. Targeting the Hippo pathway in cancer, fibrosis, wound healing and regenerative medicine. Nat Rev Drug Discov. 2020; 19(7): 480-494.
19. DeRan M, et al. Energy stress regulates hippo-YAP signaling involving AMPK-mediated regulation of angiomotin-like 1 protein. Cell Rep. 2014; 9(2): 495-503.
20. Pobbati AV, et al. A combat with the YAP/TAZ-TEAD oncoproteins for cancer therapy. Theranostics. 2020; 10(8): 3622-3635.



