
��-�����Զ�����������ϳ�;������
��-��ԭ�ϳ�;����Ϊ���������������һ�֣�O-��ԭ��ۻ�ø��Wzy��������
��;���ĵ�һ������GCL�Ͻ��йѾ����ظ���λ�ĺϳɣ�����Ҫ�����ǹѾ����ظ���λ�ڰ�������GCL-PP��ʽ�ϳɣ�Ȼ���ڧ�-��ԭ��λø��Wzx��������ת�������ʼ�϶���Խ����ظ���λ�Ķ�ۻ���Ӧ����������ԭĩ�����죬�ڷdz��ӽ�Ĥ�IJ�λ�����ۺϣ��÷�Ӧ��ҪWzy����Wzy�ĵ��ػ���������ȱʧ����ֻ���γɽ��һ���ظ���λ��LPS����ֲ��ͣ���ͨ����;���ϳɵ�O-��ԭ��Ϊ����壨���ظ���λ�ɲ�ͬ���ǻ����ɣ���
1��O-��ԭ�ظ���λ�ĺϳ�
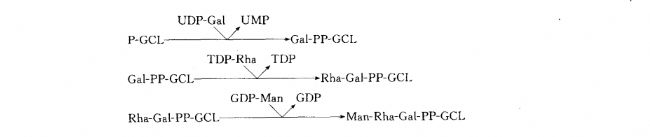
�ò���Ӧ������ϸ��Ĥ�İ����棨ͼ3-6������ɳ�ž��������ǻ����Ǽܣ������ữø��������UDP-Gal��GCL-P��Ӧ�γ�GCL-PP-Gal��UMP����ͨ�������ǻ�����Ӧ�Dz�ͬ��NDP-����ת�Ƶ�Gal-PP-GCL�ϣ��γ����ǻ����Ǽܡ�ɳ�ž���-��ԭ�����ǹǼ��ظ���λ�ĺϳ�����ͼ��
2��O-��ԭ�������ľۺ�
GCL-PP-Gal��һϵ���ǻ�ת��ø�Ĵ�������ظ���λ�ĺϳɡ��ºϳɵ�GCL-PP-�ظ���λ��Ϊһ������Ӱ�����ת�������ʼ�϶�棬��ת�˹��̿�������Wzx��������ɵģ�Wzx��һ�־���12����Ĥ�������ˮ�Ե����ʡ���Wzy���£������ۺϵ��ظ���λ����תλ���ºϳɵ�GCI-PP-�ظ���λ�ϣ�ͼ3-6������˶��������ӳ������仹ԭ�˿�ʼ�ġ�ɳ�ž�����ԭ���ǵľۺϷ�Ӧ��������ʽ��ʾ��
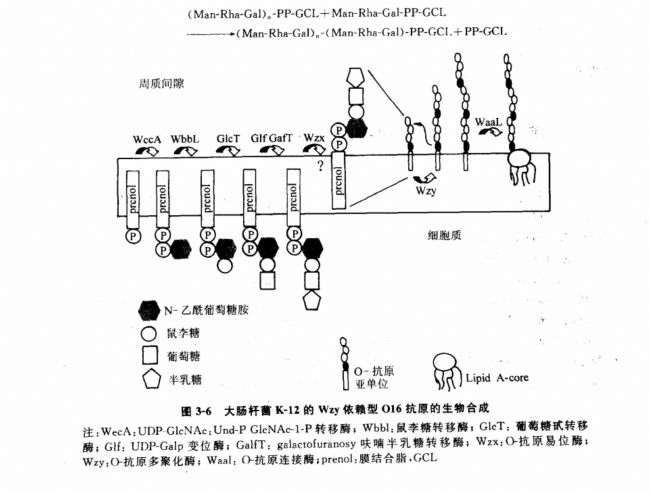
�ظ���λ�ľۺϷ�����ϸ��Ĥ�����ʼ�϶�棨ͼ3-3��ͼ3-6������תλ��������PP-GCL�ڽ�����һ���ظ���λ�ϳ�ǰ���뾭��������ø�����γ�P-GCL�����ٴν���ѭ����
3���ۺϺ������
ɳ�ž���-�����ظ���λ��ȡ���ɱ��ֳ������ԣ���Ҫ������O-��ԭ�ظ���λ�ϵ��Ǻͣ���O-������ȡ�����ŵ����ޡ��������˺�ɳ�ž���-������ǵİ�����4λ�ϱ�������ȡ�����Ӷ��γɧ�-��ԭ����122�����ڰ���������������α-1��4�������������ǡ�������ν��O-��ԭ���εĵ������ӡ��÷�Ӧ��Ҫһ��������֬�м����ʵIJ��룬��һ���ʼ�����ȷ��Ϊβ-Glc-P-GCL��
4�����ǵ�תλ
������øWaal�������£�����ԭ��ϸ��Ĥ�����ʼ�϶����core-Iipid A���ӣ��γ�������LPS���ӡ�Waal��Ŀǰ��֪��Ωһ�����ӷ�Ӧ�йص�ø�o����Ч�ذѸ߷�����������ͷ������Ѿ�������������Waal��ͬʱʶ��-��ԭ��core-Lipid A�еĺ��Ľṹ��Ŀǰ�Ը�����ø����ʶ�����١�
�˾�K-12��ijЩ��-��ԭ�ظ���λ����Ķ����������ǻ�ΪN-���������ǰ����ǰ����ǡ������ظ���λ�ĺϳ����ɶ���ù�����е�UDP-GlcNAc��GCL-P-GlcNAc-1-Pת��ø��WecA������ʼ��Ӧ�ģ�ͼ3-6�����������ظ���λ�ϳɺͶ�ۻ���Ӧ��ɳ�ž�����ͬ��

- ���������������ELISA�Լ��������ཡ������е�Ӧ��
- ʹ��Cygnus ELISA�Լ��и�������ÿհ���
- �����鷨��ҩ���ڶ��غ�������е�Ӧ��̽��
- ���������Ծ���"���ι���"�������ڶ��������ͷŻ���������ͻ��
- �˶�����Υ����̴����ئ�2�˷ܼ�����Щ����μ�⣿
- �������Լ���rCR��������C���ӣ�rFC���IJ����ԱȽ�
- ���Լ�ʹ��ָ��֮��ȷ�����������ѿ�
- ϸ���ڶ��ؼ�����ս�����ͼ��ߡ����Լ�����������Ӧ��


