
Ҷ����ӫ���Ǻ���Ǹ߷�Ӧ�����¼�����2024��4�£�
�������ǽ��ع�һ��4�·ݵ¹�WALZ����Ҷ����ӫ���Dz��뷢����7ƪ�߷����£�����Nature Communications 2ƪ��Advanced Science 1ƪ��The Plant Cell 1ƪ��The Plant Journal 1ƪ��Journal of Experimental Botany 2ƪ���¹�WALZ�����PAM����Ҷ����ӫ�����ڹ�������о�����ңң����~ңң����~
��ֲ��Ĺ�������У�ѭ�����Ӵ���(CET)��һ����Ҫ�Ĵ�л;�������ܹ��ڲ�����NADPH������£�ͨ��PSI���յ��ӣ�ά��Ҷ����Ĥ�ϵ������ݶȡ���һ���̶���ֲ����Ӧ��ͬ���ջ�����������ϵͳ���ܹ����˾�����Ҫ���塣2024��4��1�գ��ձ�������ѧ���о���Ա��The Plant Journal�Ϸ�������ΪCritical role of cyclic electron transport around photosystem I in the maintenance of photosystem I activity���о����ġ����о�ͨ�������Ͻ��е�PROTON GRADIENT REGULATION 5 (PGR5)��������ó�(KO)ͻ�����о���������PGR5��CET�еĹؼ����á�

�о��Ŷ���������CRISPR-Cas9��������������PGR5��KO��λ����ͻ���壬�ֱ�Ϊpgr5-5��pgr5-6�������ʾ��������KO��λ����ͻ������ԭ�е�pgr5-1ͻ������ȣ��ڷǹ⻯ѧ����(NPQ)���յ�������̶Ƚ��ᡣ�����PGR5������ͨ��Ӱ��NPQ�Ĵ�С������CET��Ϊ�˽�һ��̽��PGR5�Ĺ��ܣ��о���ͨ���Ŵ��������֣�pgr5-1ͻ�����д��ڵڶ���Ӱ��NPQ��С��ͻ��λ�㣬λ��pgr5-1λ������(south)Լ21 cM(centiMorgans)�������⣬�о������֣�����ʰ���-130-˿����仯��pgr5-1��λ����Ĺ�������pgr5-5�ı��ͻ����������pgr5-1ͻ���ƻ���PGR5 ���ȶ��ԣ���ͻ�䵰�ױ����˲��ֹ��ܡ���pgr5-1ͻ����ı�����Ҫ������PGR5G130S����ˮƽ�ϵ����¡��о������֣�ͬʱȱ��PGR5��NDH������(��һ��CET;��)��˫ͻ�����ڲ������ǵ�����������������������Щ˫ͻ�����PSI�����������ͣ���PSII���Խ������������CET����ά��PSI����������Ҫ��
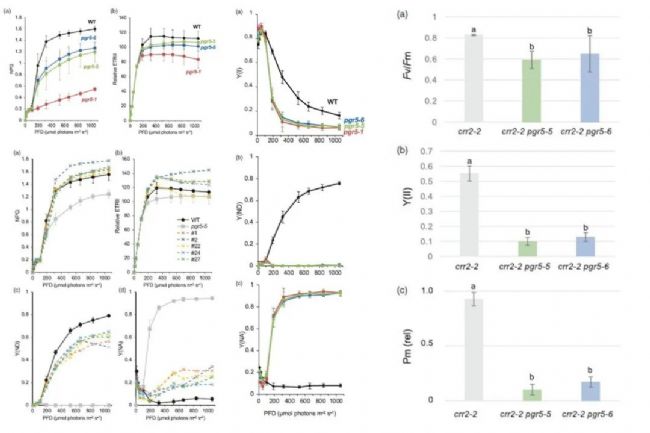
�����о�������ʾ��PGR5�ڵ���CET�еĹؼ����ã���Ϊ����ֲ�������Ӧ��ͬ���ջ����ṩ���µ��ӽǡ�ͨ����һ�����о������������������µIJ��ԣ�ͨ���Ŵ���������ǿֲ��Ĺ�����ã��Ӷ���������������Ӧ�ԡ�
��ŷ������Ҷ���ڼ��ڽ�����һֱ�����������滷���ľ�仯���ر����ڶ������¶Ⱥ��ձ仯����Ӧ����Щֲ��ͨ���ڷ���ˮƽ������ֲ������ϵ��ڹ�����÷�Ӧ����Ӧ������������ϵͳI(PSI)��ϵͳII(PSII)�Ĺ⻯ѧ���̣��Լ�������ѭ���еĶ�����̼ͬ�����ڵ�������ø���Ե���Ҷ��ҶƬ���������������£�ά�ֹ��װ�ò�ͬ���֮�������ƽ��������Ҫ������ᵼ�¹�ϵͳ���ȼ����͵��ӻ��ۣ������������˺�PSII��PSI�Ĺ����ơ�Ϊ��Ӧ�Ա����¶Ⱥ���������������ŷ��Ҷ����ֲ���ڼ��ڳ߶��϶�̬�������ϵ������������һϵ�йⱣ�����ƣ�������1.����ͳ����ķǹ⻯ѧ���(NPQr��NPQs)�Ľ�ϣ���ֹPSII���ȼ�����2.�����Ӵ����Ե�����(LEF)����·�ɵ����������(AEF)�������ӷ����������ڿ�����ѭ���ж�����̼ͬ����NADP+֮����������壻3.�ϵ�ѭ��������(CEF)����������PSI��Χѭ���ص�������(PQ)�ء�AEF��CEFͨ����ͬ�ķ���;����ֹ�����ڵ��Ӵ������л��ۣ����ڻ���Ӧ�������´ٽ��Ᵽ�����ܡ�������ǰ�Ա�ŷ��Ҷ����ֲ�^������Ӧ���о�ǿ����AEF��CEF����Ҫ�ԣ���PSI��PSII��DZ�ڶ�̬��δ�����2024��4��2�գ�����ͼ�����ѧ��Eva-Mari Aroʵ������Journal of Experimental Botany��־������ΪAccounting for photosystem I photoinhibition sheds new light on seasonal acclimation strategies of boreal conifers���о����ģ����½��Ҷ����aӫ�⡢P700����������ؼ������嵰��ȵĶ����ȼ����ֶΣ��о���Pinus sylvestris��Picea abies�ڴ���������ûָ��ڼ�PSII��PSI �Ķ�̬��ֵ��ע����ǣ����о����벢Ӧ����һ��У�����PSI���Ӳ��ʼ��㹫ʽ������չ��P700������ղ����ķ������������ڱ�ŷ��Ҷ����ʶ����PSI�����ơ�

PSI�Ĺ�������һ����Զ��������أ��йص����ױ���Ҳ�Ƚ��١��ڱ��о��У��о���Ա�ص��ע����������PSI�Ĺ����ơ�PSI�����ƻ�Ӱ�����P700����ֵ����PSI�����Ӳ��ʣ���������Ӱ������Ƿdz���̬�ģ�����ܻ���������ڱ�ŷ������Ҷ���зֱ�PSII��PSI�ڼ��ڳ߶��϶�̬�仯������������PSII��PSI���Ӳ��ʵķ���է���������ܷdz����ƣ���Ϊ���߶�����PAM�����ͱ������壬����������С�ź�ˮƽ(Fo�� Po)����̬�ź�ˮƽ(F��P)������ź�ˮƽ(Fm��Fm'��Pm��Pm')֮�������仯��Ȼ����PSII�� PSI�źŽ�������ݵ���������ԭ���ͼ���ͨ���Dz�ͬ�ġ����к���Ҫ��һ��������PSI���Ӳ��ʵ�ԭʼ����ٶ�YImaxΪ��������PSII�ĵ�Ч����ʽ(YIImax��ͨ����Ϊ Fv/Fm)�ǿɱ�ġ�
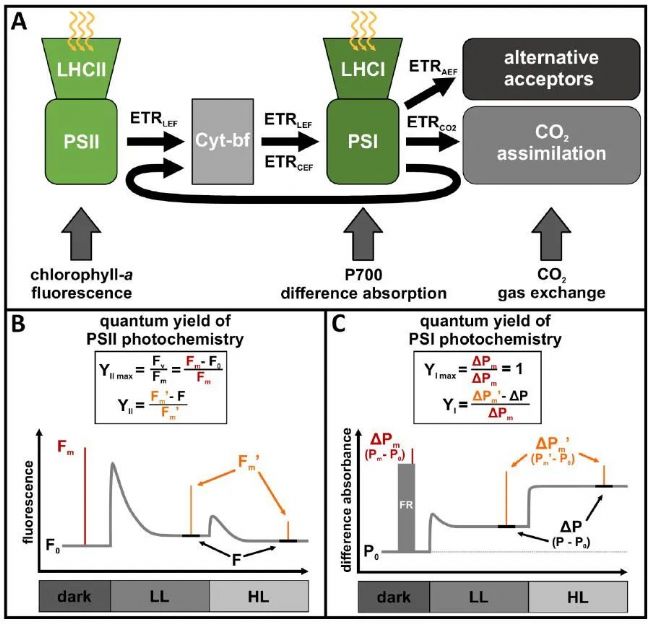
���YIֵ�㶨�ļ��費�ɱ���ػᵼ��PSI���Ӳ��ʵĶ��岻����PSI���������ã����൱���˳Ծ�����ΪPSI���������ñ��㷺��Ϊ�����������ԭ����PSI������ΔPm�Ľ���һ�¡�����PSI���������ûᵼ��PSI���Ӳ���ʧ�棬��ǰ����������ΪPSI��“©��ЧӦ”������������������ͻ������������Ƕ�PSI�ĵ��������Լ�PSI��PSII����֮���ϵ���о������������������ų�PSI�����Ƶ������£�������Խ��������Ҷ���С�
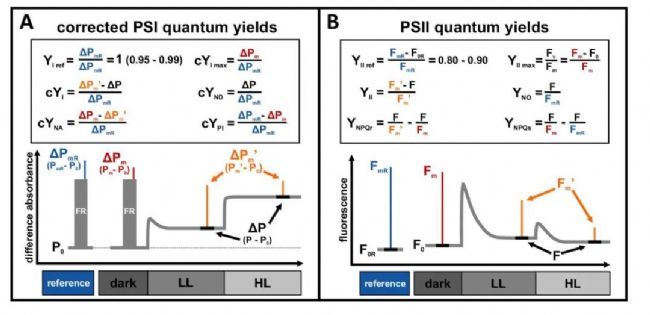
ͨ��������Ĺ�ʽ��������о�������֣�����PSI�����Ƶ���PSII��PSI���ĸ�����Ļ�ѧ���������˺ܴ�ļ����Ա仯����PSII��PSI�⻯ѧ�����ļ����Զ�̬��������ƽ�⡣ͬ�����ڿ�����PSI���������غ�֮ǰ������ѭ���������������ϵ��������ԡ���֮�����о��Ľ��ǿ���˿���PSII��PSI �Ķ�̬�Բ���Խ������ֲ�������õļ�����Ӧ�Ե���Ҫ�ԡ�������Ҷ��֮�⣬����У����PSI���Ӳ���Ϊ���ּ�ڲ���PSI��̬���ص��о��ṩ�˸���ߡ�
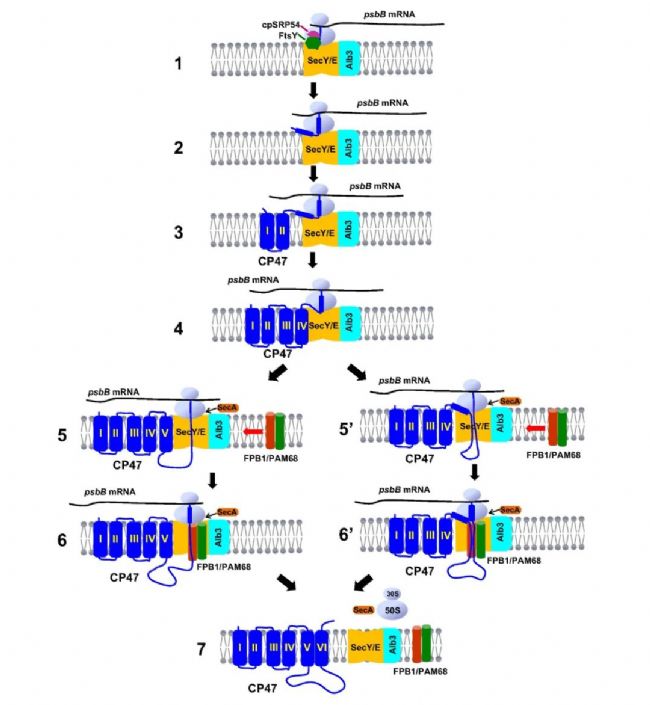
���������ֲ������������Ĺؼ����̣�����ϵͳII(PSII)����һ���̵ĺ��ġ���ϵͳII(PSII)�������ڹ������������������Ҫ�����ã�����ˮ�������������ĵ���ת�ƣ��Ӷ�����������ù����еĵ��Ӵ��ݡ�PSII����װ��һ�����ӵĹ��̣������γ�D2-Cyt b559�Ǹ����壬��ΪD1��PsbI�����帴���塣D1��ǰ��(pD1)�ڷ����������D2-Cyt b559�Ǹ����干�������ϣ��γ�PSII��Ӧ���ġ�PSII��Ӧ���ĵ���D1�������TMDs���乲�����������ѵõ������о�����D1�ĵ�һ��TMD�Ӻ�����ͨ���г���֮ǰ��������ͨ��������嵰��L420���������cpSRP54��ϣ�Ȼ��D1�ĺ�����������(RNC)ͨ��SRP����cpFtsY��������������Ĥ�е�SecY/Eתλ�ӡ�����PSII�����ǻ�D2��CP43��CP47����Ϊ�����ǹ�������װ�ģ���Ŀǰ�в����SecY/E-cpSRP54��������Ƿ�������ǵķ������װ���Լ���Щ�ǻ��Ŀ�Ĥ�ṹ�����ͨ�����Ӱ��µ������ϵ�Ĥ�С�2024��4��10�գ�Nature Communications���߷����Ϻ�ʦ����ѧ����ΰ����ʵ������ΪThylakoid protein FPB1 synergistically cooperates with PAM68 to promote CP47 biogenesis and Photosystem II assembly���о����ģ����½�ʾ��һ���µĵ�����FPB1(Facilitator of PsbB biogenesis1)������PSII����װ�а�������Ҫ��ɫ������PSII����������ġ�

�о���Աͨ���Ŵ�������ʵ�鷢�֣�FPB1����֪��PAM68����Эͬ���ã���ͬ�ٽ���CP47—PSII�����ǻ�������ϳɡ���ȱ��FPB1��PAM68������£�CP47�ĺϳ����ʽ��ͣ������γɹ����Ե�PSII�����壬����ֲ����������Ч�Ĺ�����á������о������֣�FPB1��һ��λ��Ҷ������Ĥ�ĵ����ʣ�����PAM68��SecY/Eתλ�Ӻ�Alb3����ø����ã�����Э��CP47�ڷ��������˳��Ƕ��Ҷ������Ĥ�����⣬ͨ�������������������о���Ա�۲쵽��û��FPB1��PAM68������£��������ڷ���CP47ʱ�������Ե���ͣ������������������ڴٽ�CP47�Ĺ�������������Źؼ����á���ͼΪ���о������Ҷ������CP47�Ĺ�������װ�Լ�FPB1��PAM68���ܵĿ���ģ�͡�
A hemoprotein with a zinc-mirror heme site ties heme availability to carbon metabolism in cyanobacteria (Nature Communications, IF=16.6)
�����Ƕ����������ѧѭ������Ҫ�����ߣ���������ռ�������������� 25%��Ϊ�˷�����Щ���ܣ����������������ͨ����������ϸ�������ұ���ȷ������������(��Ѫ����)������ϳ���������ƥ�䣬�������ںϳɹ�Ϻͺ������ø����塣�����ڵ��Ӵ����з�����Ҫ�����⣬����������(��Ѫ����)���������ϸ�����ܡ����磬Ѫ������һ�ֹؼ����źŷ��ӣ��ɵ��ڴ�ת¼���źŴ����Լ���Ҫ�����ʷ������صȸ���ϸ�����̡���ˣ�����������ڽ��������漰Ѫ���صĵ��ڻ��ƣ���ά����ѵĹ��Ч�ʣ�ͬʱ��������Ӧ�������ˡ���½��ֲ���У���������һ������Ѫ���صĻ��������ƹȰ���-tRNA��ԭø(GluTR1)�ķ�ȣ����ֵ���������������������ϳɵĵ�һ����GluTR1�ķ������ڶ��ڷ�ֹѪ���ؼ����м����Ĺ�������������Ҫ����������Ѫ���صĸ��������������� GluTR1 ��ϵ���(GBP)������GluTR1�ĵ��ڽṹ���Ϻɷ�ֹGluTR1��Clp����ø���⡣Ѫ������GBP��ϻ�������������ã�����GluTR1���⡣GBP��һ��N �˷ֲ�Ͱ״�ṹ��(���߰���������ø�������һ����)��һ������δ֪��C�˽ṹ��DUF2470��ɡ��������ṹ��Ҳ�����ڲ���ϸ��Ѫ���ؽ��������ȡ��HugZ���������С�DUF2470�벻ͬ�ķ��ӹ����йأ��ýṹ����Ϊ����������HugZ��Ѫ���ؽ�Ͽڴ�������GBP �У�DUF2470�����ʶ��GluTR1����ȻѪ������GBP�Ľ���ѱ�֤ʵ����Ѫ���ؽ����ʽ��δ�ӽṹ��ȷ��������Ŀǰ����ȷ���������ṹ������һ�������Ѫ���ء���ˣ�DUF2470�Ľṹ-���ܹ�ϵ��δȷ����4��12�գ�Nature Communications���߷���������Դ�����ϻ������о���Crysten E. Blaby-Haasʵ������ΪA hemoprotein with a zinc-mirror heme site ties heme availability to carbon metabolism in cyanobacteria���о����ġ������о��Ŷ�ͨ�����ϵͳ����������ѧ�����ͽṹ������������һ��������Ѫ���ص����壬���а���һ������δ֪��DUF2470�ṹ��Ѫ������������п��ϵ��鰱��л�������λ���γ�һ�����Ե�˫�ضԳ�п-�鰱��-��-�鰱��-пλ�㡣��Ͻṹ���������������ʵ�飬�о���Ա��һ��֤����Dri1��Ѫ���ؽ��������۰���������������ø�ķ�������֮����ڹ�����ϵ������������úͺ���֮������������Ե��ڹ�ϵ����������������ԭ���������ձ���ں���ͬԴ�ṹ���Ѫ���ش�л�ĵ��ף��о���Ա���DRI��������صĽṹ��;��ǰ��ΪDUF2470���ڷ���ˮƽ����ΪѪ���������Ե��ڽṹ�����á�

���⣬����ϸ���о��������Dri1�ܹ����Ѫ���أ������侧��ṹ��ʾ��һ��п��Ѫ����λ�㡣���ֽ����ͨ��п���Ӻ��ض����鰱��л��鵼�ġ�Dri1������������ø(succinate dehydrogenase, SDH)�ķ��������йأ���ʾ��һ���������ԵĹ�����úͺ�������֮��ĵ�����ϵ��Dri1�Ľṹ��صĽ������Dri1��û��Ѫ����ʱΪ���壬�����Ѫ���غ��γɶ����塣ͨ��X���߾���ѧ��С�ͽ�X����ɢ��(SAXS)������Dri1���������Ľṹ���о������Dri1��ΪѪ���������Ժ�ת¼�������ӵ�ģ�ͣ�����ͨ��ֱ�ӻ���Ӱ������������ø������Ļ�����������Ӵ������ĵ��ء�Dri1������ϸ���е����ð����Թ�����úͺ������õĵ�������ֱ��Ӱ�����Ӱ�죬�Լ��������������¶�ϸ��������Ӱ�졣Dri1����������ϸ���еľ��Ա����Ա������õ�����Ѫ������̬�з����Źؼ��ұ��ص����á�

Unique photosynthetic strategies employed by closely related Breviolum minutum strains under rapid short-term cumulative heat stress (Journal of Experimental Botany, IF=6.9)
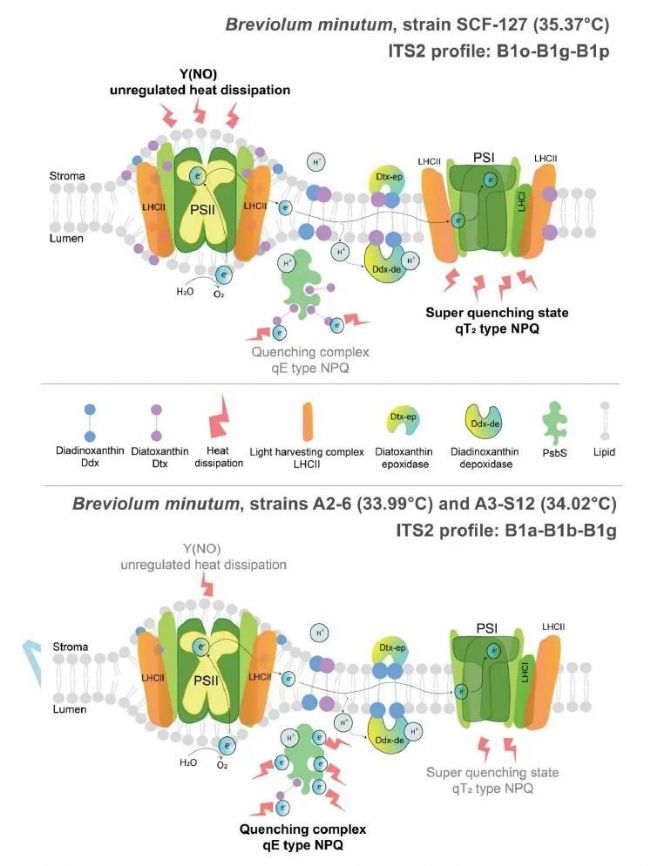
����������ɺ���ͺ�����һϵ�д̰����������İ��ڹ����壬�ṩ��Ϲ�̼�Ի�ȡ�����ṩ�ĵ�����Щ�����в�ȵ�����ˮƽ��������������������Ҫ�����������ļ������ڼ䣬���������ᵼ�¹�������ɺ������֮�������Ⱥ��ʧ������ɰ��������磬Durusdinium���е�����ͨ����Cladocopium���е����ָ����������ߵİ������ԣ�Cladocopium C15�����в�ͬ��Ա�Ĵ�����ɺ����ͬ�����������йء���������ֲ���������ȡ������������в���±��ֹ��ϵͳII������ϲ�����(Fv/Fm of PSII)��ˮƽ̼�̶���������2024��4��18�գ�Journal of Experimental Botany��־�����߷���ī������ѧ�����ѧѧԺΪ������λ������ΪUnique photosynthetic strategies employed by closely related Breviolum minutum strains under rapid short-term cumulative heat stress���о����ġ�

���о����µĺ��������ǹ�������������ص�Breviolum minutum�����ڿ��ٶ����ۻ���Ӧ��(26-40°C)�������õĶ��ع�����ò��ԡ��о��Ƚ�����Щ������������ԡ��⻯ѧ�ͷǹ⻯ѧ���𡢹��ɫ�ص��ѻ�����״̬�Լ�����������(ROS)�Ļ��ۡ���Ҫ���ֺͽ��۰�������ͬ��B. minutum��������˲�ͬ�ĹⱣ�����ԣ����������Ǹ��Ե��ϲ��������Բ�ͬ���о��ṩ��֮ǰδ֪���������������Ᵽ������֮���������ԣ�����������������ͨ���ǹ�����Ŀ��ٷ��ɺ�״̬ת����ֵĺ�ɢ֮�����ƽ�⡣�����������Ե�B. minutum����(B1-B1o-B1g-B1p)���ֳ���ǿ���ѻ��������ã���������Ĥ���۵�ǿ����أ�������ͨ��Ĥ�ӹ̽�����������С�������о�������������������ص�B. minutum������֧���������ԵĹⱣ�����ơ��о���̽��������Ӧ���£���ͬB. minutum����Ĺ�����ò����仯��������ϵͳII�������Ч��(Fv/Fm)��ʵ�����Ӳ���(ΔF/Fm′)���ǹ⻯ѧ����(NPQ)�Լ�ROS�IJ�����ͨ�����ɷַ�����PCA���Ͳ�ͬ��ͳ�Ʋ��ԣ��о���ʾ�˲�ͬ��������Ӧ���µĹ�������Ӧ���졣���»�����������Ӧ���£�B. minutum����ĹⱣ�����ƵĶ�̬�仯���������ٷ��ɵ�NPQ(qE��)�������ɵ�NPQ(qI��)���Լ�״̬ת��(qT1��qT2��)�����о��Ľ����֮ǰ�۲쵽�����ǵ�����Exaiptasia diaphana����Ư��������һ�£�ǿ��������Կ��ٶ�����Ӧ��ʱ����ͬ�����ȡ�˲�ͬ�Ĺ�����ò��ԡ�
Rewiring photosynthesis by water-soluble fullerene derivatives for solar-powered electricity generation (Advanced Science, IF=15.1)
������ý�����ת��Ϊ��ѧ������Ȼ��������Ҫ������ת�����̡��ⷴӦͨ���������ѽ�ˮ��������������̬���ӣ�����ϵ��Ӵ��������ɻ�ԭ��NADPH��ATP�����ڰ���Ӧ�����̼��Ӧ����������Ԫ�����˹�Ԫ���ı��ϵ��Ӵ��ݷ����������������ת��Ч�ʣ������������������������졢��ϲ����ϵͳ��Ȼ������Ȼ���ϵͳ��ϸ����л���طdz����ӣ���ͳ���﹤�̲�������ʵ�ֹ�ϵ��Ӵ��ݹ��̵ĸ���͵��ء�2024��4��22�գ��й���ѧԺ�����о��������Ŷӵ��컪ΰ��ʿ���������ѧ���ĺ����о�Ա��������Advanced Science��־�Ϸ�����ΪRewiring photosynthesis by water-soluble fullerene derivatives for solar-powered electricity generation���о����ġ����������һ�ָı��ϵ��Ӵ��ݷ�����²��ԣ���������ά̼���ײ��ϴӹ�ϵ��Ӵ������н������Ӳ����ݵ����⣬�Ӷ����������ת��Ч�ʡ��о���Ա��ƺϳ���һ�ֱ��������ɵ�ˮ���Ը���ϩ���ײ��ϣ������ײ��ϱ�����ϸ��������Ըı��ϵ��Ӵ��ݷ��ٽ���ϵ���������������������ܶ����һ����������

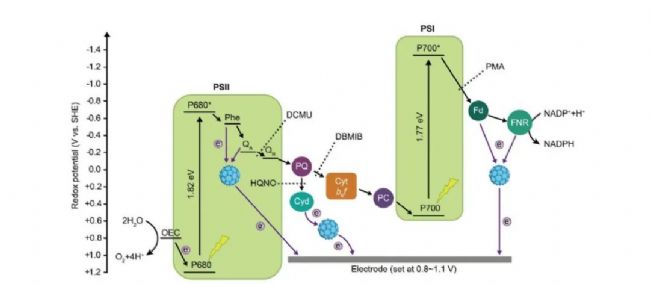
��ô������ϩ����������δٽ�����ĵ������Ź����أ��о���Ա���������Ե��Ӵ������Ƽ����з�������������������ϸ���У���ϵ�����Ҫ�ӹ�ϵͳ I ���Σ��ر��Ǵ��������ش������������⡣������ϸ�����ո���ϩ�������ϵ�������Ҫ�ӹ�ϵͳ I ���Σ��ر�����������ԭ���� Fd ����������������ϩ�ڹ�ϵ��Ӵ������ϵ�����λ����Ҫλ�ڹ�ϵͳ I ����ࡣ����˲̬���չ�����̬ӫ���������һ����ʾ��������ϩ���������ϵͳII��ϵͳI�ĹⷴӦ���ľ�����ֱ������á����˸���ϩ������ C60-DMePyI ֮�⣬�о���Ա���������������ֲ�ͬ���ܻ�����ϩ���ײ��϶Թ����������Ӱ�죬�Լ���Щ���ײ��ϵ� Zeta ��λ�p����Ч�ʺ�������ԭ���ԡ��������������ϩ�ı��ϵ��Ӵ��ݷ����������Ҫȡ����������ɺ�������ԭ���ԡ�������ɵĸ���ϩ������������������ϸ��Ĥ����á�ͬʱ�����ײ�����ˮ����ϵ�еĿ���������ԭ�������䷢�ӵ������幦�ܵĹؼ����ء��ۺ���Щ������������������ɵ�ˮ���Ը���ϩ�������ܹ�������ϸ����ϵ��Ӵ������е�������ԭ���ķ�������ã��ı��ϵ��ӵĴ��ݷ���ʹ��ϵ��Ӵ�����ϸ����������Ӷ��������ϸ���Ĺ���������о�����������������ʶ����ϸ���й�ϵ��Ӵ��ݵ����ϣ�ҲΪ����ά�������帻��ϩ���ײ�������������ת���е�Ӧ���ṩ�˿��ܡ�
Photosynthetic control at the cytochrome b6f complex (The Plant Cell, IF=11.6)
�������������պ����ù��ܣ����Dz��ܴ�����ܣ�������DZ��������ƽ����Щ��л������ɺ����ġ�����ⷴӦ��CBBѭ�������ʲ�ƥ�䣬���չ�ǿʱ���γɻ�����(ROS)����ϸ�����ˣ���������ʱ��ᵼ��ϸ�������½����������ޡ����ڹⷴӦ��һ����Ҫ�����ǹ�Ͽ���(PCON)������NADPH��ATP�����ɳ���CO2�̶�����������£�ͨ��ά�ָ�ΔpH�Ե���-����ת�Ʒ�Ӧ���з������ڡ�PCONͨ�����ڵ��Ӵ������ʣ�������ϵͳI(PSI)���ܹ��յ����ڲ������ջ����н��е�ʵ��֤����PCON��ֲ�������ͻָ������ı�Ҫ������ȱ���ù��̵�ͻ���������PSI�������ƻ���
PCON��һ�ַ����Ա������ƣ���ͨ��ά�����Ե��Ӵ���(LET)������NADPH��ATP�������̼(CO2)�̶���Ӧ��������֮���ƽ�⣬��Ч�����˹��ܶԹ�ϵͳI(PSI)��DZ������ֲ���������ھ����������¶���Ҫ���ȿ�����α��� PSI�ģ���Ϊֲ��ȱ��ר�����PS��Ŀ�����ѭ��������ζ���κ����˶��ᵼ�³�ʱ��Ĺ����ƺ�����˥�ˡ�LET��CO2�̶���Ӧ֮��IJ�ƽ���ͨ����������ĤΔpHˮƽ����֪�������չ���ʱ��ΔpH�����ߡ�PCON�ĵ��ͻ���������ͨ��ΔpH��ϸ��ɫ��b6f�����Ե��Ӵ���(LET)��������(PQH2)����������еķ������ء�PCONͨ���˻��Ʊ����˹�ϵͳI (PSI)���������Ҷ����(P700)������״̬����ʹ����CO2�̶���Ӧ��δʹ�õĶ�������ܹ�ͨ��������鱻��ȫ�ش���2024��4��26�գ�Ӣ��л�ƶ��´�ѧMatthew P Johnsonʵ���ҵ�Gustaf E Degen��The Plant Cell��־������ΪPhotosynthetic control at the cytochrome b6f complex���о����������£������Ա���ֲ��Ϊ�о�����̽����PSI��������������β����ģ�PSI�����ƶԹ�����ú�������Ӱ�죬���������˽�PCON���ڷ�������½�չ�����չ����δ�������������� PCON��߹��Ч�ʵ�ǰ����
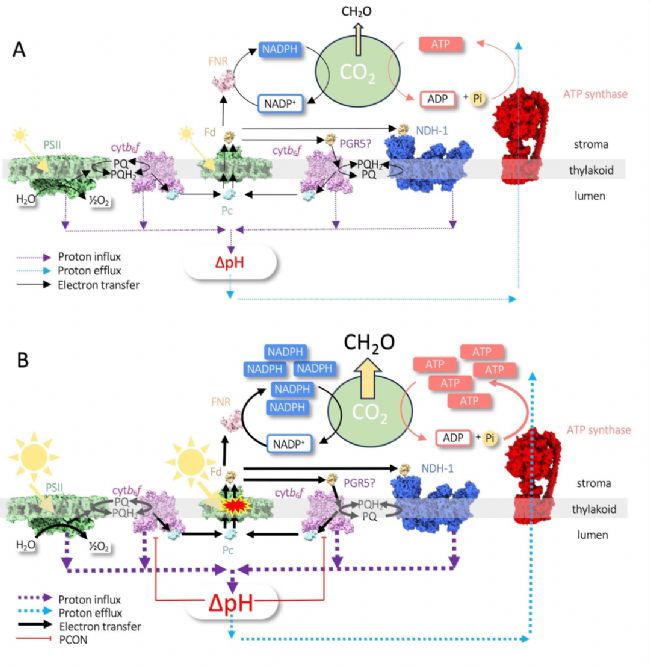
��������ΔpH��qE��ͬ���䶯̬�仯�Թ�ǿ�ȱ仯���ֳ��ͺ�ЧӦ��P700����������ȴ�ܼ���Ѹ�ٵط��ɡ�ʵ���ϣ������Ͻ�ֲ����ʹ�ÿ��������Ҳ����ջ��Ʊ����� qE��������������ȷ�ظ��ٹ���ǿ�ȵı仯����PCON����Ӧ��Ҫǿ�öࡣ�������ͨ������qE�ķ������������õظ��ٹ���ǿ�ȱ仯���Ӷ��ɹ�������������ת��������P700������˵�Dz���Ҫ�ġ�Ȼ�����ڹ�β��(Setaria viridis)�����Ͻ��У�ISP�ǻ��Ĺ�����Ӷ����ϸ��ɫ��b6f�������ˮƽ���ѱ�֤������ǿ������������������CO2ͬ�����ã��������������PSI�ĵ�����������ǿ������á���ʵ�ϣ���β��ISP��������ϵ��ʾ����Ұ�����ߵ�PSI�������͵�PSI����ˮƽ���������ٶȸ��졣ͬ������������ϸ��ɫ��c6��ϸ��ɫ��b6f��PSI֮��Pc������������壬Ҳ��������������ˣ����µ���ϸ��ɫ��b6f��ΔpH�������ԣ�����ʹ��ϵͳ�Ե��������������ͣ�����������ε��ӻ�IJٿ�Эͬ���ã�����ͨ��������CBBѭ���е�����ø����������������
P700����(Y(ND))���ڱ���PSI�����Ƶ���Ҫ�����Զ����ġ�Ȼ������ͳ�����ϵĹ�����(��ϸ��ɫ��b6f�Ե���������������ΔpH�仯)�������������õģ���ΪKP700��P700red t½ͨ�����������ǿ�ȵ����Ӷ��仯���෴�����������Ǻ㶨�ģ����Y(ND)��ӳ��Cytb6f��LET ���ʵĹ������ƣ�������������û�иı䡣��ͻ����PCON��Y(ND)֮�������������ˣ�Ҫȷ��PCON�ı仯����ҪPSI���ٻ�ԭ��(P700red t½)����������ȷ��˵��Ҫȷ��Cytb6f�����ı仯��Ӧ����Cyt fredt½��Pc��ԭ��˥��(Pcredt½)��ͨ����Щ����ȷ�IJ����������Թ۲쵽�����ΔpH�½�����ֵ����(����pgr5������������ֲ���й۲쵽������)��PCON�ͻ��½������ϸ��ɫ��b6f ISP�����仯��PCON�ͻ����ӡ�Ȼ�����ڵ�CO2��в�������£�������ֵΔpH�⣬����Ҫ�����������յ�PCON���Ѿ�����˼���������ԭ�ͻ��ڽṹ��PCON������ڻ��ƣ�����Ӧ�ý�һ������ʵ����֤��
�����
1. Kobayashi, R., et al. (2024). "Critical role of cyclic electron transport around photosystem I in the maintenance of photosystem I activity." The Plant Journal n/a(n/a).[DUAL-PAM-100]
2. Grebe, S., et al. (2024). "Accounting for photosystem I photoinhibition sheds new light on seasonal acclimation strategies of boreal conifers." Journal of Experimental Botany. [GFS -3000 & DUAL-PAM-100]
3. Zhang, L., et al. (2024). "Thylakoid protein FPB1 synergistically cooperates with PAM68 to promote CP47 biogenesis and Photosystem II assembly." Nature communications 15(1): 3122. [IMAGING-PAM & PAM-2500 & DUAL-PAM-100]
4. Grosjean, N., et al. (2024). "A hemoprotein with a zinc-mirror heme site ties heme availability to carbon metabolism in cyanobacteria." Nature communications 15(1): 3167. [DUAL-PAM-100]
5. Deore, P., et al. (2024). "Unique photosynthetic strategies employed by closely related Breviolum minutum strains under rapid short-term cumulative heat stress." Journal of Experimental Botany. [IMAGING-PAM]
6. Zhu, H., et al. (2024). "Rewiring Photosynthesis by Water-Soluble Fullerene Derivatives for Solar-Powered Electricity Generation." Advanced Science n/a(n/a): 2310245. [DUAL-PAM-100]
7. Degen, G. E. and M. P. Johnson (2024). "Photosynthetic control at the cytochrome b6f complex." The Plant Cell. [DUAL-KLAS-NIR & DUAL-PAM-100 &P515/535]

�绰��021-32555118




