
Dickkopf-1在循环拉伸期间上调UHRF1促进平滑肌细胞增殖和迁移
心血管系统不断地暴露在由血流和压力决定的各种机械力之下。有充分证据表明,血压升高(高血压)会导致血管重塑。血管壁介质中的血管平滑肌细胞 (VSMCs) 是维持血管稳态所必需的,在体内受到机械循环拉伸。随着病理性拉伸的增加,VSMCs从收缩型转变为合成型,其特征是增殖和迁移活动增加,收缩性丧失,细胞外基质产生异常。然而,机械循环拉伸调节 VSMCs 功能的分子机制仍有待进一步阐明。
DKK1 (dickkopf-1) 是 Dickkopf 家族中研究的最深入的分泌蛋白。越来越多的研究表明,DKK1具有肿瘤促进作用。近年来,DKK1也被确立为心血管疾病的新媒介。此外,DKK1 也可作为预测急性缺血性中风和急性冠状动脉综合征患者临床结果的潜在生物标志物。其他一些先前的研究表明, DKK1 在调节人类血管平滑肌细胞的功能中的作用相互矛盾。因此,探索DKK1 在VSMCs 中的作用相关的分子信号通路仍然是一个关键的挑战。
由山东大学齐鲁医院、中国医学科学院、心血管重塑与功能研究教育部重点实验室等单位的专家学者先前已经证明,DKK1参与了动脉粥样硬化的发展。靶向DKK1疗法可能不仅有抗肿瘤作用,而且对心血管疾病也有保护作用,一举两得。因此,揭示DKK1在心血管疾病发展中的作用,对于指导针对DKK1的抗肿瘤药物在心血管疾病患者肿瘤中的应用具有重要意义。
该团队已经证实,DKK1在内皮细胞中的表达在体内扰动流和体外振荡剪切应力 (OSS) 处理下上调。DKK1 的敲低减弱了OSS 诱导的单核细胞粘附和内皮损伤。目前尚不清楚 DKK1 是否通过机械拉伸调节平滑肌细胞的功能。
因此,该团队进行了深入研究,于 International Journal of Biological Sciences 上联合发表了题为《Dickkopf-1 promotes Vascular Smooth Muscle Cell proliferation and migration through upregulating UHRF1 during Cyclic Stretch application》的研究成果。在这项研究中,以人主动脉平滑肌细胞(HASMCs)和小鼠为实验对象,探讨了DKK1在病理牵张条件下动脉重塑发展中的作用及其机制。
实验结果:
机械过度拉伸增加了 VSMCs 中 DKK1 的表达
腹主动脉缩窄 (AAC) 小鼠的收缩压 (SBP) 和舒张压 (DBP) 显著高于假手术对照组小鼠。如图1 A、C,在手术后1 周,与假手术对照组小鼠相比,AAC 小鼠的胸主动脉和冠状动脉中膜中检测到 DKK1 蛋白水平显著增加。取胸主动脉并通过蛋白质印迹分析,在AAC小鼠中DKK1的相对蛋白表达水平显著增加(图1 D)。通过ELISA测定小鼠血清中的DKK1水平,两组之间未观察到血清DKK1 水平的差异。
对人 VSMCs 进行了进一步的体外实验,以验证 DKK1 响应高水平拉伸的变化。使用循环拉伸加载系统,对 HASMCs 进行不同时间长度(0、3、6、12 或 24 小时)的高强度拉伸(18%),HASMCs 和培养上清液都表现出 DKK1 蛋白水平随时间依赖性增加 (图1 E、F),而当细胞处于5% 循环拉伸时没有观察到变化。将HASMCs 在无拉伸、正常拉伸 (5%) 或高强度拉伸 (18%) 的情况下处理 6 小时,然后收集细胞裂解物用于蛋白质印迹分析。DKK1 表达在高水平拉伸处理的细胞中显著高(P < 0.05,图1 G)。
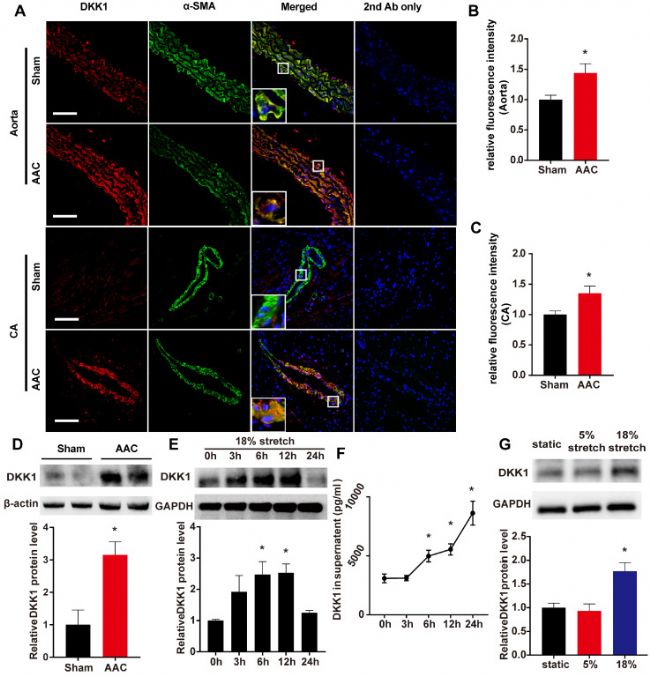
图 1
DKK1 有助于在机械拉伸下调节 VSMC 功能
DKK1 在调节 VSMC 功能调控中的作用目前尚不清楚。实验通过针对DKK1 的小干扰 RNA (siRNA) 转染到细胞中来降低 VSMCs 中 DKK1 的蛋白质表达。与用干扰siRNA转染相比,用DKK1-siRNA 转染 24 小时有效地抑制了 DKK1 mRNA 水平(图2 A)。
在用 DKK1-siRNA 或干扰 siRNA 转染后,HASMCs 用 5% 或 18% 的拉伸刺激 24 小时,然后进行 EdU 和 CCK-8 检测细胞增殖,Transwell 检测细胞迁移。EdU 和 CCK-8 检测表明,18% 的循环拉伸显著促进了 HASMCs 的增殖,而敲低 DKK1 抑制了这种作用(P<0.05,图2 B-D)。Transwell 测定结果表明,DKK1-siRNA 组中迁移到下腔室的细胞数量显著低于对照组 (图2 E、F)。
通过WB法分析全细胞裂解物的增殖标记物PCNA和收缩表型标记物α-SMA的蛋白质水平。与正常拉伸(5%)处理相比,高水平拉伸(18%)上调了PCNA 的蛋白质水平并下调了 α-SMA 的蛋白质水平(图2 G-I)。通过敲低 DKK1 的表达,这些变化被部分逆转。外源性rhDKK1的添加促进了细胞增殖和迁移,促进了PCNA的表达并抑制了α-SMA的表达(图2 J-Q)。
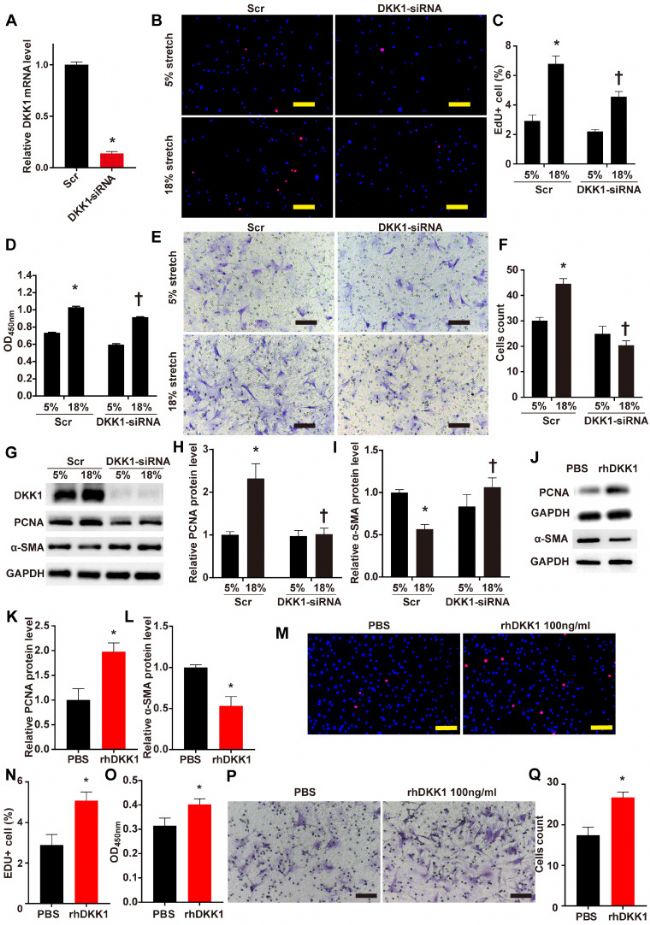
图 2
DKK1 通过靶向 UHRF1 参与调控 HASMC 功能
考虑到 DKK1 被称为典型 Wnt 信号通路的拮抗剂,研究人员进行了实验以确定在循环拉伸应用过程中,典型的Wnt通路是否参与了DKK1在HASMC 中的作用。
在差异表达的基因中,重点研究了一个特定的基因,UHRF1。最近的研究结果表明,UHRF1 可能在调节血管平滑肌细胞的增殖、迁移和表型方面发挥重要作用。实验验证了在高水平拉伸下UHRF1的蛋白质水平显著增加,而在正常条件下没有检测到变化。当 DKK1 在高水平拉伸下被 siRNA 敲低时,UHRF1 表达在 mRNA 和蛋白质水平上显著下调(P < 0.05,图3 C-E)。此外,rhDKK1 的刺激显著增加了 HASMCs 中 UHRF1 的表达 (图3 F)。
为了进一步研究 UHRF1 是否介导 DKK1 对 HASMCs 的影响,用 siRNA 敲低 UHRF1 48 小时,并在 rhDKK1 刺激的情况下用正常 (5%) 或高水平拉伸 (18%) 处理细胞 24 小时。然后通过EdU 和 CCK-8 分析评估细胞增殖,Transwell 分析评估细胞迁移, qRT-PCR 评估 RNA 的干扰效率(P < 0.05,图3 G)。在 rhDKK1 刺激的 HASMCs 中,敲低 UHRF1 显著减弱了细胞增殖和迁移的增加(P<0.05,图3 H-L)。同样,蛋白质印迹分析表明,敲低 UHRF1 后,PCNA 的蛋白水平显著降低,α-SMA 的蛋白水平升高(P<0.05,图3 M-O)。
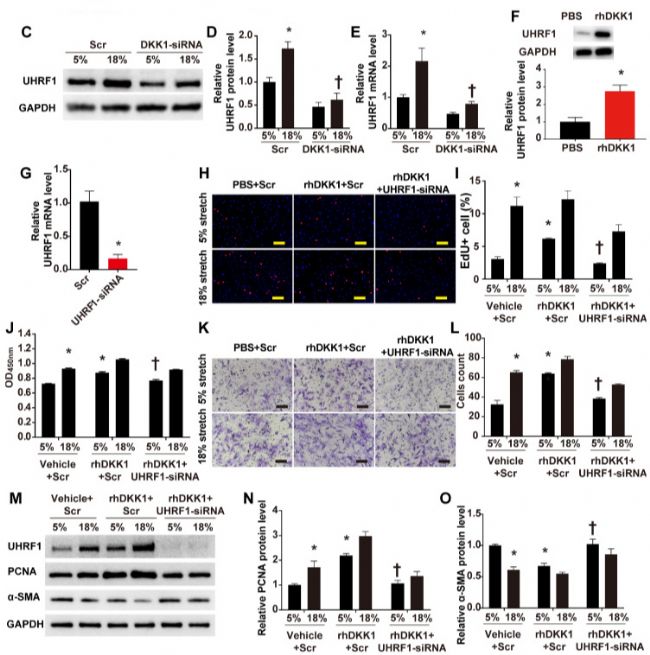
图 3
此外,实验还证实小鼠平滑肌 (SM) 特异性 DKK1 缺失可改善血管重塑,说明DKK1 参与了由高水平拉伸引起的血管重塑和 VSMC 功能的改变;并且DKK1 是通过 YAP-TEAD 通路调节UHRF1。
实验结论:
在这里,实验证明 DKK1 是机械拉伸诱导的 VSMC 表型转换的关键介质。数据表明,高水平的拉伸可在体内和体外诱导 DKK1 的表达。在体内,实验发现 SMCs 中DKK1 基因的缺失减轻了 AAC 诱导的血管重塑。在体外,机械拉伸诱导的DKK1 通过YAP-TEAD 通路上调 UHRF1,并导致 VSMC 增殖和迁移的增强。
总之,该研究表明,DKK1 通过 YAP-TEAD 通路调节 UHRF1 表达,从而介导平滑肌细胞功能的机械拉伸调控。
参考文献:Zheng TF, Liu XL, Li X, Wang QQ, Zhao YC, Li X, Li MM, Zhang Y, Zhang M, Zhang WC, Zhang C, Zhang Y, Zhang M. Dickkopf-1 promotes Vascular Smooth Muscle Cell proliferation and migration through upregulating UHRF1 during Cyclic Stretch application. Int J Biol Sci. 2021 Mar 21;17(5):1234-1249. doi: 10.7150/ijbs.56247. PMID: 33867842; PMCID: PMC8040467.
小编旨在分享、学习、交流生物科学等领域的研究进展。如有侵权或引文不当请联系小编修正。微信搜索公众号“Naturethink”,学习更多关于流体机械力学刺激细胞培养相关知识吧!



