
EPAS1 ͨ����Ƥ֬������ȡ����Ѫ����������������Ӳ���ķ���
EPAS1 Attenuates Atherosclerosis Initiation at Disturbed Flow Sites Through Endothelial Fatty Acid Uptake
Keywords: atherosclerosis; diet, high-fat; endothelial cells; obesity; plaque, atherosclerotic.
��������Ӳ���߿鷢չ���ȶ����ڱ�¶��Ѫ�����ҵĶ�������ѻ�����ЩѪ������ѧ���������ͷ��Ⱥ�����Ļ�е�ڼ���Ӧ�����յ��ֲ���Ƥϸ����ECs�������ϰ����Ӷ������߿��γɡ�ȫ�����еķ������л�쳣�йأ�����Ѫ֬�쳣��Ѫ�ǣ���Щ���Ƕ�������Ӳ�����������ء�Ѫ�����һ����� EC ��Ѫ֬�쳣��Ѫ�ǵ������ԣ�Ȼ����������ϵ����ķ��ӻ�����Ȼ֪֮���١�
��Ƥ��̬��ά�������ڵ��ڴ�лƽ���ת¼����֮����۸��ӵ�����á�ֵ��ע����ǣ�ȱ���յ����ӣ�HIF��ת¼���� HIF1A �� HIF2A��EPAS1���ڸõ������������ź������á�EPAS1 ��Ѫ������ѧ�еĹ����о����١���֪ EPAS1 ͨ���ȶ�����Ѫ����������������������ٽ�Ѫ���γɡ�EPAS1 ���������������������̣�������̥��������ϸ�����ɡ��������������л��Ȼ����EPAS1 �ڶ�����̬�� EC ��е��Ӧ�Ͷ�������Ӳ���е�DZ�������Բ������
�����Ӣ��л�ƶ��´�ѧת����ѧ�о������ص۹�����ѧԺ�����ķ��о�������ʿ��������ѧҽԺ�ٴ���ѧ�о����������Ŷ��о��� EPAS1 �ڶ�������Ӳ����չ�е����ã��ر�����Ѫ�����ҵ������о�֤�� EPAS1 ����Щ������ͨ���ٽ�֬�����л����ǿ��Ƥ��̬�����ƶ�������Ӳ�������� EPAS1 ȷ��Ϊ��������Ƥ�Լ���Ӧ����Ӧ֮��Ĺؼ��м价�ڣ���ʾ�˷��֡�Ѫ�����ҺͶ�������Ӳ����ϵ�ĸ��ӷ��ӻ��ơ��о��ɹ������� Circulation Research �ڿ���Ϊ“EPAS1 Attenuates Atherosclerosis Initiation at Disturbed Flow Sites Through Endothelial Fatty Acid Uptake”��

���ȣ�Ϊ�˷��� EPAS1 �Ƿ��Ѫ������Ӧ��ʹ������ƽ�а�ϵͳ����������Ѫ����Ƥϸ����PAEC����¶�ڲ�ͬ����Ӧ����������72 h������Ӧ����HSS��13 dynes/cm2�����ͼ���Ӧ����LSS��4 dynes/cm2���������Ӧ����LOSS��4 dynes/cm2��1 Hz��˫������ӡ����ʾ����LSS����HSS ��ȣ���¶�� LOSS�� EPAS1 �������ӣ�ͼ1 A����Ȼ������С���������� EC �е� EPAS1 ˮƽ������������ȣ������ʱ�¶����Խϵ͵�ʱ��-ƽ�� WSS ���Ⱥ������������ LOSS����ͼ1 B�����������о����һ�£�С������������ӫ��Ⱦɫ��ʾ���� HSS ������ȣ�EPAS1 �� LOSS ���������ʣ�����������ͼ1 C����LOSS λ��� EPAS1 һ���ֶ�λ��ϸ���ˣ��������ǻ�Ծ�ģ�ͼ1 C������ˣ�EPAS1 �����ⱻ LOSS ��ǿ�����ڱ�¶�� LOSS ��С���������Ķ�������Ӳ��������
Ϊ���������ֶ���������Ƥ EPAS1 ˮƽ��Ӱ�죬�� WT С��¶�ڸ�֬��������ʳ���� 22 �ܡ����������ı���Ⱦɫ��ʾ��������ι���Ķ���С����ȣ���¶�� HFD ��С�� LOSS ����� EPAS1 ˮƽ�������͡���ˣ��������ص� HFD ι���ή�� LOSS ��������Ӳ��������� EPAS1 ���
����о���Ա��ͼȷ�������� EPAS1 ���͵Ļ��ƣ����ص��ע�˷����д�л�仯�� EPAS1 �����Ӱ�졣ͨ����С��ע����������������STZ���յ�������ģ�ͣ�����Ⱦɫ��ʾ����STZ������С���У������� LOSS ����� EPAS1 ����û�иı䣬������Ѫ�Dz��������� EPAS1��
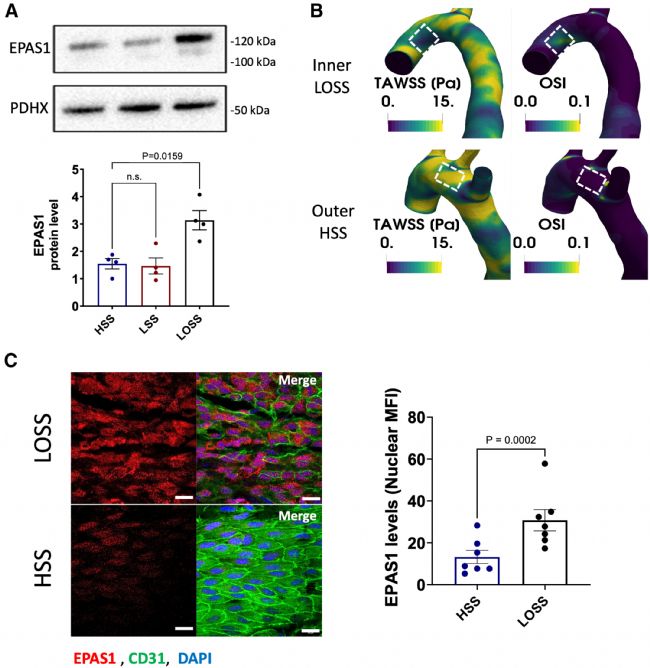 ͼ1 EPAS1�ڱ�¶�ڵͼ�Ӧ������Ӧ���µĶ�������Ӳ�����Բ�λ������
ͼ1 EPAS1�ڱ�¶�ڵͼ�Ӧ������Ӧ���µĶ�������Ӳ�����Բ�λ������
��������ͨ�����ܲ����أ�Sulforaphane����������С����������֬Ѫ֢�� EPAS1 �����е�DZ�����á��ܲ�������֪���Ըı�֬���ͱ�����Ƥ������������ܲ��������������˱�¶�� HFD ����С��Ѫ���и���������TGs��ˮƽ��ͼ2 A������Ѫ�ǣ�ͼ2 B�������غ�Ѫ���ܵ��̴�ˮƽ��HDL�����ܶ�֬���ף��� LDL/VLDL�����ܶ�֬����/�����ܶ�֬���ף�ˮƽû�������ı䡣�ܲ����ش������ָ��� EPAS1 �� LOSS ����ı�����ʶ���������� HSS ����ı��ͼ2 C����
�� SFN �����ķ���С���У�EPAS1 �Ļָ�������Ѫ�� TG ������أ���ˣ����� TGs ���ܲ�������� EPAS1 �����ƣ����� LOSS �������� PAEC ��¶������֬���ᣨFFAs����������֤������ӡ����ʾ����¶�����ᣨOA��������ᣨPA���� EPAS1 ˮƽ�������͡�Ϊ���о�DZ�ڻ��ƣ����� EPAS1 �� PHD2 �� PHD3 �����н��⡣PA �� OA �������� PHD2 ˮƽ������ FFAs ����ͨ���յ� PHD2 ���� EPAS1��ͨ������������Ⱦɫ�õ���һ����֤����Ƿ���С����ȣ�ιʳ HFD �ķ���С���� LOSS ����� PHD2 ������ǿ��
�ܲ����ؿ���ͨ����������� NRF2 �����ƻ������� EC ����ֱ��Ӱ�졣��ˣ��ڴ��ڻ����ܲ����ػ������� NAC ���������PA�� OA ���� PAEC ���о���ͨ·�� EPAS1 ����е�DZ�ڲ��롣�ܲ����غ� NAC �������� PA �� OA �������� PAEC �е�ƽ�� EPAS1 ˮƽ������û�дﵽͳ��ѧ���壬�������ų� SFN ͨ�����ٻ���������� FFA �鵼�� EPAS1 ���ƵĿ����ԡ����⣬�û��ƿ����漰���� PHD2���䱻 SFN �� NAC ���͡���ˣ��ܲ����ش����ķ���С���� EPAS1 �Ļָ������漰 SFN ����Ƥ��ֱ�����á���Щ�۲���������EPAS1 �ڷ����еĽ��Ϳ��������ڸ߸�������Ѫ֢-FFA ��л��ϸ������Ӧ���ľֲ��ı����ϣ�������Ƥ PHD2 ���յ���
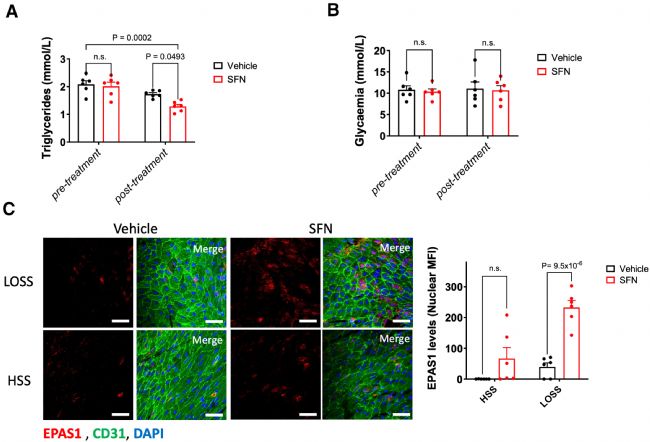 ͼ2 �ܲ����ؿ���ȷ���С��� EPAS1ˮƽ��
ͼ2 �ܲ����ؿ���ȷ���С��� EPAS1ˮƽ��
��һ���أ����������Ƿ������Ѫ�������Ѫ��ϵͳ�й۲쵽�����е� EPAS1 ���ƣ�����С����ֵ���Ѫ����Ƥϸ���� EPAS1 ���ͣ�ͬʱѭ��Ѫ�� EPAS1 Ҳ���١�
ͨ���ȽϷ��ֺͷǷ��ָ���� PHD-EPAS1 ͨ·�������о�������ٴ�����ԡ�ELISA �����ʾ��EPAS1 �����зǷ��ֶ��յ�Ѫ�������о��ɼ������ڷ��ָ�����ȱʧ���������١�Ȼ���о����ٴ����ֻ���Ѫ�� EPAS1 �����Ƿ���ѭ��֬�ʺ� ROS �ı��йء���Ƿ��ָ�����ȣ������ߵ� FFAs ˮƽ�������ӣ���ȴû�дﵽͳ��ѧ���塣������ͷǷ�����֮��Ѫ����ܿ������������ơ����ֻ��ߺͷǷ��ֶ�����Ƥ��֬���з����Ѫ�� EC �� EPAS1��PHD2 �� PHD3 ת¼��Ķ���ʵʱ PCR ��ʾ�����������ȣ����ָ���� PHD2 �� PHD3 ˮƽ�������ӣ��� EPAS1 mRNA ����û�иı䡣��Щ���ݱ�����EPAS1 ����Ѫ֬�쳣�Լ� EC �� PHD2 �� PHD3 ����������صķ��ָ������ܵ����ƣ���һ�۲������ڷ���С��ģ���еĹ۲���һ�¡�
Ϊ�˷��� Epas1 �ڶ�������Ӳ���е����ã����� Epas1EC-KO �� Epas1EC-WT����С��¶�� HFD �Բ����ߵ��̴�Ѫ֢��ͼ3 A������ƤEpas1 ȱʧ��Ѫ�����̴�ˮƽû��Ӱ�죨ͼ3 B����Ȼ����С���������еĶ�������Ӳ���߿����Ը���ͼ3 C����������������Ҳ�����˰߿���γɣ�֤ʵ�����������ȣ�Epas1EC-KO С����ڸ���İ߿飨ͼ3 D������Щ���ݱ�������Ƥ EPAS1 �ɷ�ֹ��������Ӳ����
Ϊ���˽���Ƥ EPAS1 �Ĺ��ܣ���Epas1EC-KO�Ͷ���С����������н����˵�ϸ��RNAseq�������� 6 ����ͬ��ϸ���أ�����Epas1 Ӱ�� EC �����ԡ�����ע����ʾ��Epas1 ���صĴظ����˶��ֲ�ͬ�Ļ������������ϸ�����ź�ͨ·��ת¼/RNA �ӹ���ϸ����ֳ�ʹ�л����Epas1EC-KO��Epas1EC-WTС��ϸ�����н�һ���Ĺ���ע�ͣ������˶�������Ļ�������������뼡�����ס��߶������Ĥ�����롢ת�ˡ��������ữ��������ͷ�����MAP ��ø�źŴ������ǵ��ס���Щ���ݿ������ɼ��� EPAS1 �����ڶ�����Ƥϸ�����������в����л���ڡ�
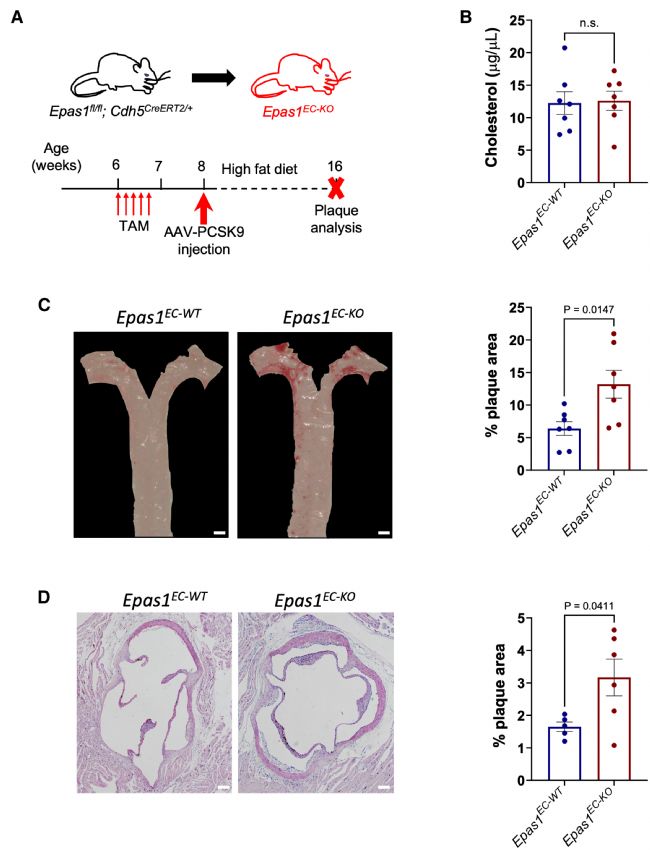 ͼ3 ��Ƥ Epas1 �ɷ�ֹ��������Ӳ����
ͼ3 ��Ƥ Epas1 �ɷ�ֹ��������Ӳ����
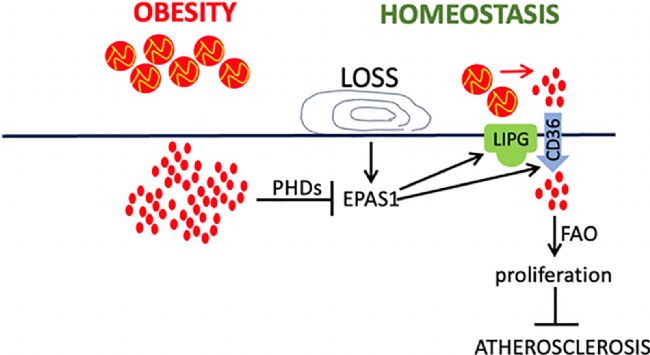
����� LOSS ������������ PAEC ���õ�EPAS1���о������֬�����л��DZ�����á��ݹ۲죬EPAS1 ��Ĭ���±�¶�� LOSS �� EC �Ļ��������ʽ���Լ80%��ͼ4 A�����������ֺ�����ǿ��������֬����β��������Щ���ݱ�����EPAS1 ��֬����β-����-�����Ժ��������������ӡ���һ���о� EPAS1 �ں����е����ã�����EPAS1 ͨ������֬����β���������ں��������Ҷ���������Ա�����Ӱ����Խ�С��
ֵ��ע����ǣ�EPAS1 ��Ĭϸ���к������ļ��ٿ���ͨ��������Դ�� OA ����ȣ�ͼ4 A��������� EPAS1 ����ͨ������ EC �������� FFAs������������֬���� β-����;����������ʵʱ PCR ������ʾ��EPAS1 ��Ĭ������mRNA ˮƽ�Ͻ�����Ƥ֬��ø��LIPG���ͷֻ���ԭ��36��CD36���ı��ͼ4 B��C����
Ϊ��ȷ�� Epas1 �� CD36 �� LIPG ��Ӱ�죬��С�������������˱���Ⱦɫ��WT С����ʾ���� HSS ������ȣ�CD36 �� LIPG �� LOSS ��������������ͼ4 D��E�������������ȣ�Epas1EC-KO��LOSS����CD36��LIPG�ı����������ͣ�ͼ4 D��E��������Epas1��LOSS���������ַ��Ӹ����ı�Ҫ������Ϊ������ LOSS ���� CD36 �� LIPG �ľֲ������Ƿ���֬�ʴ�л������أ�ͨ���ڷ�����ͽ�С��¶�ڼ��Ը�֬Ѫ֢������Ѫ��TGˮƽ��90 ���Ӻ��������ӡ�ֵ��ע����ǣ��� HSS ������ȣ��������� LOSS �����֬�λ�����ǿ�������ֲ�֬����ȡ��ǿ����Щ���ݱ�����EPAS1 ͨ���������Э�� FFA ������ CD36 �� LIPG �ı���Ƕ�������Ӳ�� LOSS ������Ƥ֬����β-�����DZ���ġ�
���� HIF1A ���� LOSS ������ǽͽ⣬��ˣ������� EPAS1 �Ը�ͨ·��DZ�ڹ��ס�EPAS1 �ij�Ĭʹ�ǽͽ���ǽͽ��������͡�HK2 �� PFKFB3 ���� HIF1A ���ڵĹؼ��ǽͽ���ڼ�������ӡ����ʾ��EPAS1 �ij�Ĭ����Ӱ��������ˣ�EPAS1 ͨ��һ�ֲ�ͬ�� HIF1A �Ļ��ƶ��ǽͽ��������ʶȵ�Ӱ�졣
��Ƥ֬����-β������DNA ����ϸ����ֳ������أ���ˣ�ʵ����� EPAS1 ������ͨ��֬�����л��֧����Ƥ��ֳ�����������ȣ�Epas1EC-KOС���EC��ֳ���Լ��٣��������Ƥϸ��Epas1��EC��ֳ������ġ�ͬ�����Ա�¶�� LOSS �� PAEC �ķ���������EPAS1 ��Ĭ����ֳ�������١�
Ϊ�˵��� EPAS1 ��֬����β-��������ֳ��Ӱ�죬ͨ������Դ�� OA ���� EPAS1 ��Ĭ�� PAEC �����֬�����л���ָ���л���������� EPAS1 ��Ĭ�� PAEC �� EC ��ֳ��Ƶ�ʣ����� EPAS1 ͨ����ǿ֬�����л��������ֳ����Щ������ʾ��EPAS1 ͨ������ EC ��л��̬����ֳ��������Ѫ���ܵ����ŵ����Ӷ�������Ӳ���������á�
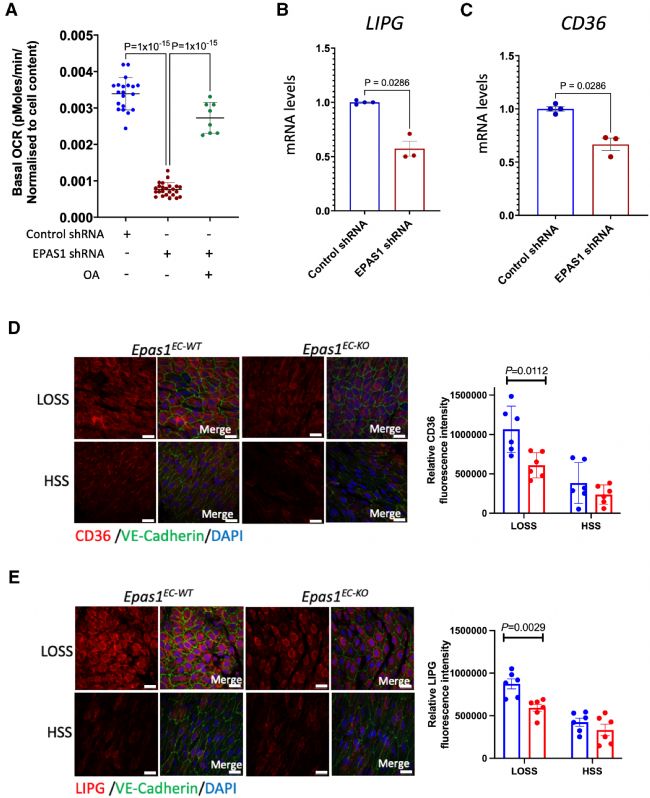 ͼ4 ��Ƥ EPAS1 ͨ�� LIPG �� CD36 �������֬�����л��
ͼ4 ��Ƥ EPAS1 ͨ�� LIPG �� CD36 �������֬�����л��
 ͼ5 ͼ�θ�Ҫ
ͼ5 ͼ�θ�Ҫ
��Ƥϸ�� EPAS1 ͨ��֬������ȡ�ʹ�лά�� EC ��ֳ���Ӷ�����Ѫ����������Ķ�������Ӳ����������Ƥ��;���ڷ������ܵ����ƣ�����һ���µĸ�������-PHD2 ����;�������� EPAS1 ���
��֮�����о����ǿ���� EPAS1 �ڶ�������Ӳ������������ȱ����ͨ���ٽ�֬�����л��֧����Ƥ����������������Ӳ�������ֱ�����;���ܵ����ֵ�Ӱ�죬��ʾ��һ�ֽ�ϵͳ��Ѫ�ܷ����� EC �����ϰ���ϵ���������ͷ�����ϵ����Щ���ֶ�����Ѫ�ܷ��յķ��ָ�����ٴ���������DZ�����塣
�ο����ף�Pirri D, Tian S, Tardajos-Ayllon B, Irving SE, Donati F, Allen SP, Mammoto T, Vilahur G, Kabir L, Bennett J, Rasool Y, Pericleous C, Mazzei G, McAllan L, Scott WR, Koestler T, Zingg U, Birdsey GM, Miller CL, Schenkel T, Chambers EV, Dunning MJ, Serbanovic-Canic J, Botrè F, Mammoto A, Xu S, Osto E, Han W, Fragiadaki M, Evans PC. EPAS1 Attenuates Atherosclerosis Initiation at Disturbed Flow Sites Through Endothelial Fatty Acid Uptake. Circ Res. 2024 Sep 27;135(8):822-837. doi: 10.1161/CIRCRESAHA.123.324054. Epub 2024 Sep 5. PMID: 39234692; PMCID: PMC11424061.
ԭ�����ӣ�https://pubmed.ncbi.nlm.nih.gov/39234692/
IF��16.5
0009-7330 (Print-ISSN)
1524-4571(Online-ISSN)
ͼƬ��Դ�� ����ͼƬ����Դ�ڲο�����
С��ּ�ڷ�����ѧϰ�����������ѧ��������о���չ��������Ȩ�����IJ�������ϵС�������������κε��뷨�Լ����飬��ӭ��ϵС�ࡣ��л��λ������Լ���ע���������www.naturethink.com���ע“Naturethink”���ںţ��˽����������ݡ�
����˽⣺���������ϵͳ



