
文献解读:苜蓿单核转录组揭示时空共生知觉和早期反应

期刊:nature plants
影响因子:15.8
主要技术:植物单细胞抽核测序
导语
豆科植物根瘤菌共生关系的建立需要时间和细胞类型特异性地精确协调复杂的反应。遇到根瘤菌后,宿主植物中基因表达水平在最初几小时内发生快速变化,为植物关闭防御并与微生物形成共生关系做好准备。在这里,我们应用单核 RNA 测序技术对根瘤菌处理后30分钟、6小时和24 小时的苜蓿根系进行了表征。我们发现 30 分钟时表皮和皮层发生了剧烈的全局基因表达重编程,这些变化中的大多数在6小时内恢复。此外,在非浓缩细胞因子处理后,植物防御反应基因在30分钟内被激活,随后在6小时内被抑制。我们发现,在接种点头因子后30分钟,招募根瘤菌所需的黄酮合成酶基因在皮层细胞中高度表达,而在其他类型的细胞中则没有。共生固氮基因富集的基因模块结果表明,MtFER(MtFERONIA)和LYK3(LysM域受体样激酶3)对共生信号具有相似的响应。我们进一步发现,MtFER可以被LYK3磷酸化,并参与根瘤菌共生。我们的研究结果扩大了我们在单细胞水平上对动态时空共生反应的理解。
技术服务
植物单细胞抽核测序
研究结果
1. snRNA-seq揭示了苜蓿根的细胞异质性
为了分析早期反应的转录组重编程,我们在7日龄的树干树幼苗上接种了NFs。然后收集处理后0.5h(hpt)、6 hpt和24 hpt的根系敏感区(NFs),并接种缓冲结瘤培养基(BNM)作为模拟对照,制备snRNA-seq文库(图1a)。我们在4个文库中共获得了25276个高质量的单核转录组,覆盖30469个基因,每个核的中位基因为1018个,每个核的中位唯一分子标识符(UMIs)为1390个(补充数据1)。整合scVI数据后,鉴定出10个细胞簇(图1b和补充图1)。然而,我们发现在统一流形近似和投影(UMAP)图中,簇0和簇3难以区分;即使是聚类0中最特异性表达的基因在聚类3中也倾向于上调(补充图1),这导致我们合并了这两个细胞簇,从而得到了9个细胞簇(图1c)。对于聚类注释,我们使用了之前在M. truncatula中描述的15个细胞类型特异性标记(补充表1),我们能够将大部分聚类分配到一个特定的细胞类型(图1d和扩展数据图1)。
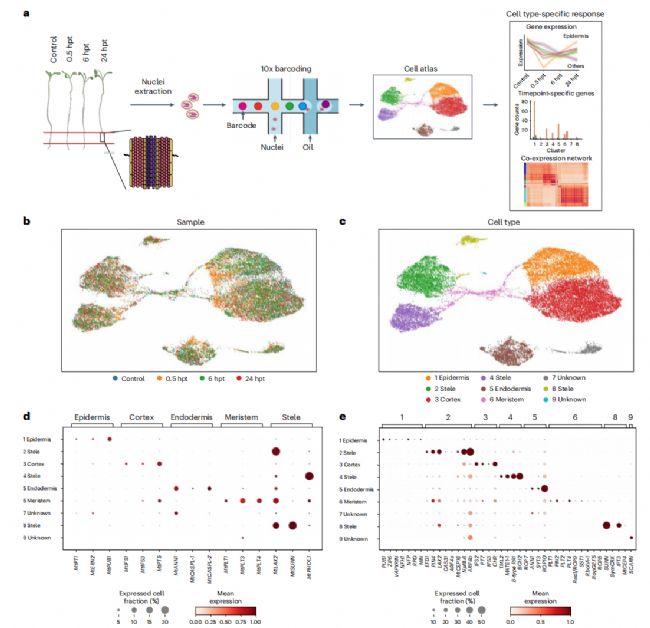
图 1
2. 广泛表达的基因反应模式的分析
为了研究早期信号通路中基因的反应模式,我们根据每个细胞簇中不同时间点的表达情况,将4个时间点的1868个基因分为6个基因类别(图2a和补充数据3)。共有838个不能准确地分配到这六个类别的基因被丢弃(方法)。从第2类到第6类的5个基因类别在所有细胞簇中都表现出相似的表达模式(图2b)。接下来,我们研究了每种细胞类型对NFs的反应。通过整合四个snRNA数据集从不同的时间点,与上述结果一致,我们发现转录组在0.5 hpt显示最低的相似性与其他时间点,表明根经历了广泛的转录组重编程迅速NFs刺激和逐步恢复后6hpt(图. 2c和补充图.6)。之前的全根研究21也揭示了根瘤菌接种后1h的瞬时重编程,我们的结果表明,瞬时重编程在表皮和皮质细胞中最为明显,突出了这两种细胞类型在NFs感知中的重要作用(补充图6)。我们还发现,在0.5 hpt时的时间点特异性上调基因数量最高,证实了0.5 hpt在早期反应中的唯一性(图2d和补充数据4)。我们通过添加一个新的生物复制来验证了这些发现,并通过使用完全基于组合的方法(方法)分析两个重复的数据集,获得了高度一致的结果(扩展数据图4。值得注意的是,对这些诱导基因的基因功能富集分析显示,在0.5 hpt时,几乎所有分化的细胞类型(除中柱亚型簇8外)都诱导了植物防御相关通路,特别是在表皮和皮质中。在6 hpt时,这些细胞在表皮中仍然活跃(图2e和补充图7–9)。
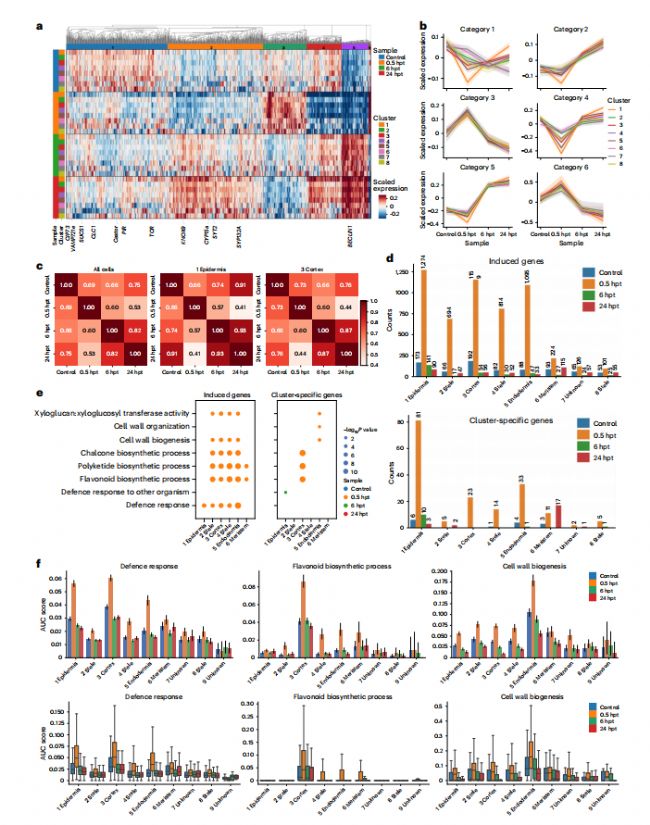
图 2
3. 表皮细胞的一种亚型是SNF所必需的
通过重新聚类,表皮可进一步分为两种亚细胞类型(亚簇1-0和1-1亚簇)(图3a)。在所有四个样本中都发现了这两个子簇,这表明在应用NFs之前,差异导致了两个子簇的差异(图3b)。此外,我们的列表中 1-1specifically 表达的基因 GH9CI的 GUS 信号主要标记成熟区的根毛和表皮细胞(图3c),这与拟南芥根毛和表皮中根毛标记基因 AtGH9Cl的表达模式一致。与根毛在根瘤菌感染初期的重要作用一致,我们发现已知SNF 基因在子集群 1-1中的比例显著高于 1-0(图 3d、e和补充数据2)。此外,与丛枝菌根共生(AMS)相关的GO功能也被富集(图3f)。
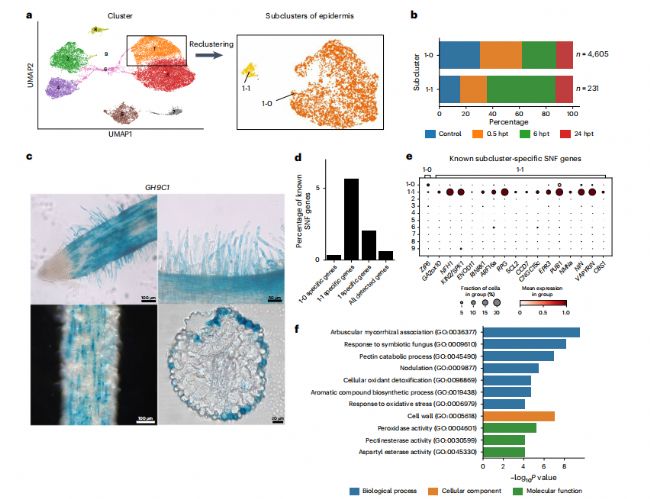
图 3
4. MtFER与LYK3共表达
为了进一步探索它们的功能,我们在0.5hpt时提取了表皮和皮质细胞,以构建共表达网络(图4a和补充数据7)。结瘤所需的乙烯响应因子2(ERN2),PUB1和LYK3图4a、c)MtFER(Medtr7g073660),拟南芥铁藻的明显同源物(扩展数据图6),也在LYK3的模块中被发现。LYK3可以磷酸化PUB1,并在早期反应中调节结瘤。此外,我们发现这三个基因的表达水平均在0.5 hpt时达到峰值,随后逐渐下降,这表明MtFER可能与PUB1和LYK3发生相互作用(图4d)。
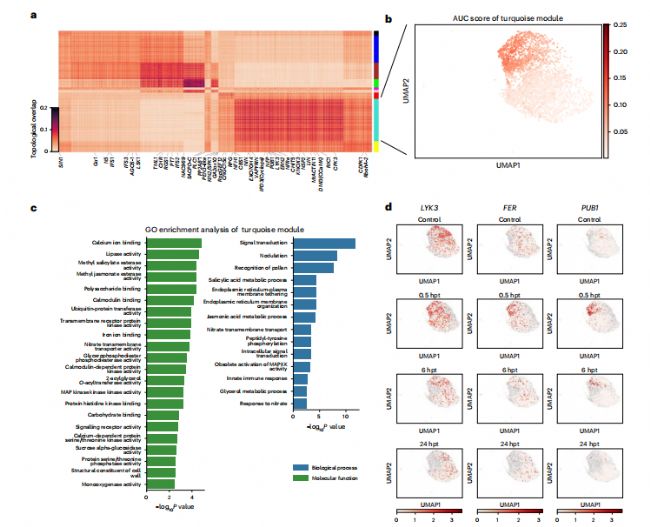
图 4
5. MtFER的表达模式与LYK3相似
我们注意到pMtFER:GUS在根尖高表达,而pLYK3::GUS在该区域弱表达(图5a和扩展数据图7)。有趣的是,我们的数据显示,在酵母双杂交(Y2H)实验中,MtFER蛋白可以与LYK3相互作用(图5b)。当LYK3-FLAG和MtFER-HA在本烟草叶片中共表达时,我们通过共免疫沉淀(Co-IP)进一步证实了相互作用(图5c)。最后,我们在体外激酶实验中发现LYK3可以磷酸化MtFER(图5d)。我们的研究结果表明,根瘤菌共生过程中对MtFER的需求可能与其与LYK3的相互作用有关。
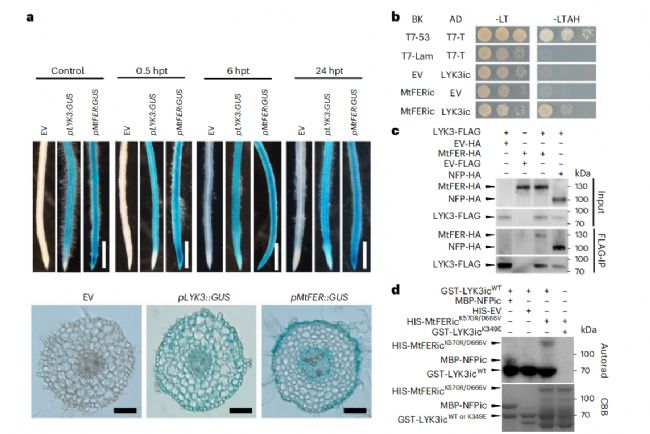
图 5
参考文献:
[1]Liu Z , Yang J ,Long, YanpingZhang, ChiWang, DapengZhang, XiaoweiDong, WentaoZhao, LiLiu, ChengwuZhai, JixianWang, Ertao.Single-nucleus transcriptomes reveal spatiotemporal symbiotic perception and early response in Medicago[J].Nature Plants, 2023, 9(10):1734.



